

MtcB, a member of the MttB superfamily from the human gut acetogen Eubacterium limosum, is a cobalamin-dependent carnitine demethylase
- PMID: 32571881
- PMCID: PMC7443480
- DOI: 10.1074/jbc.RA120.012934
MtcB, a member of the MttB superfamily from the human gut acetogen Eubacterium limosum, is a cobalamin-dependent carnitine demethylase
Abstract
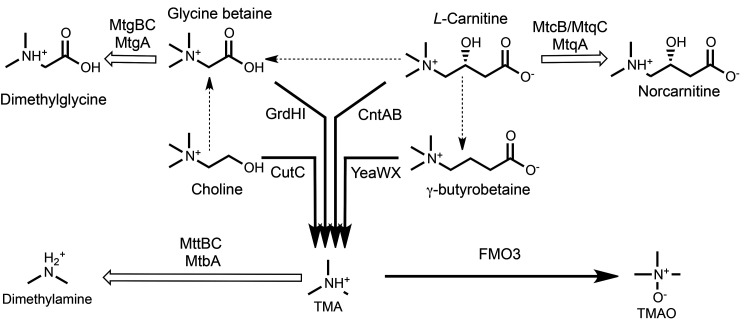
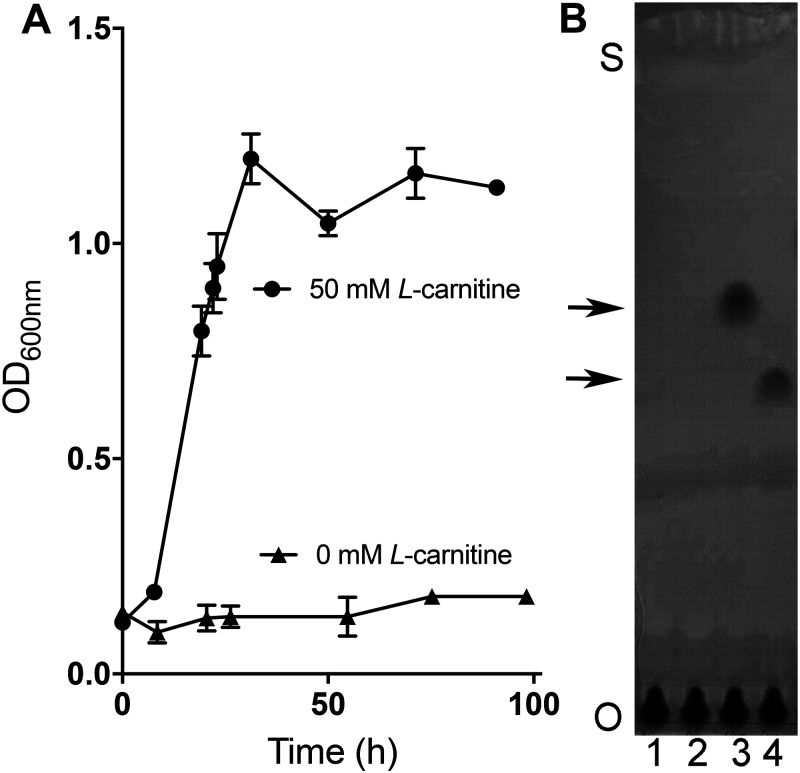
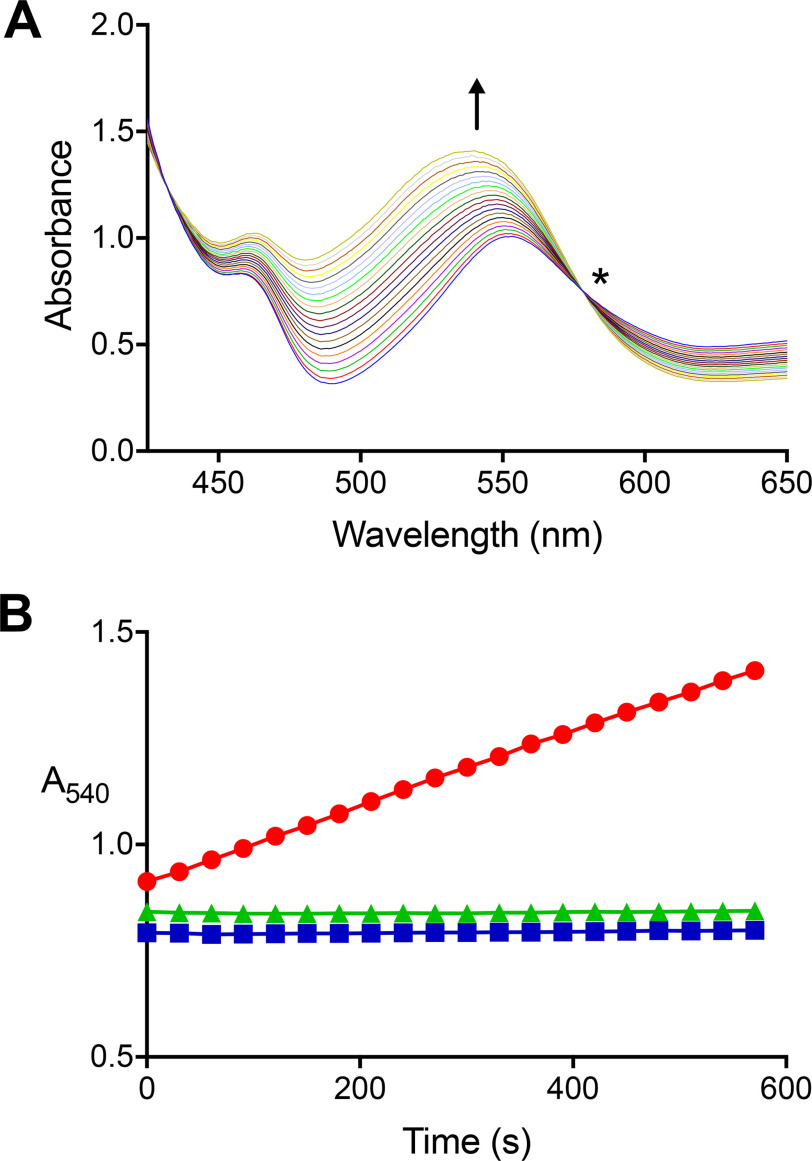
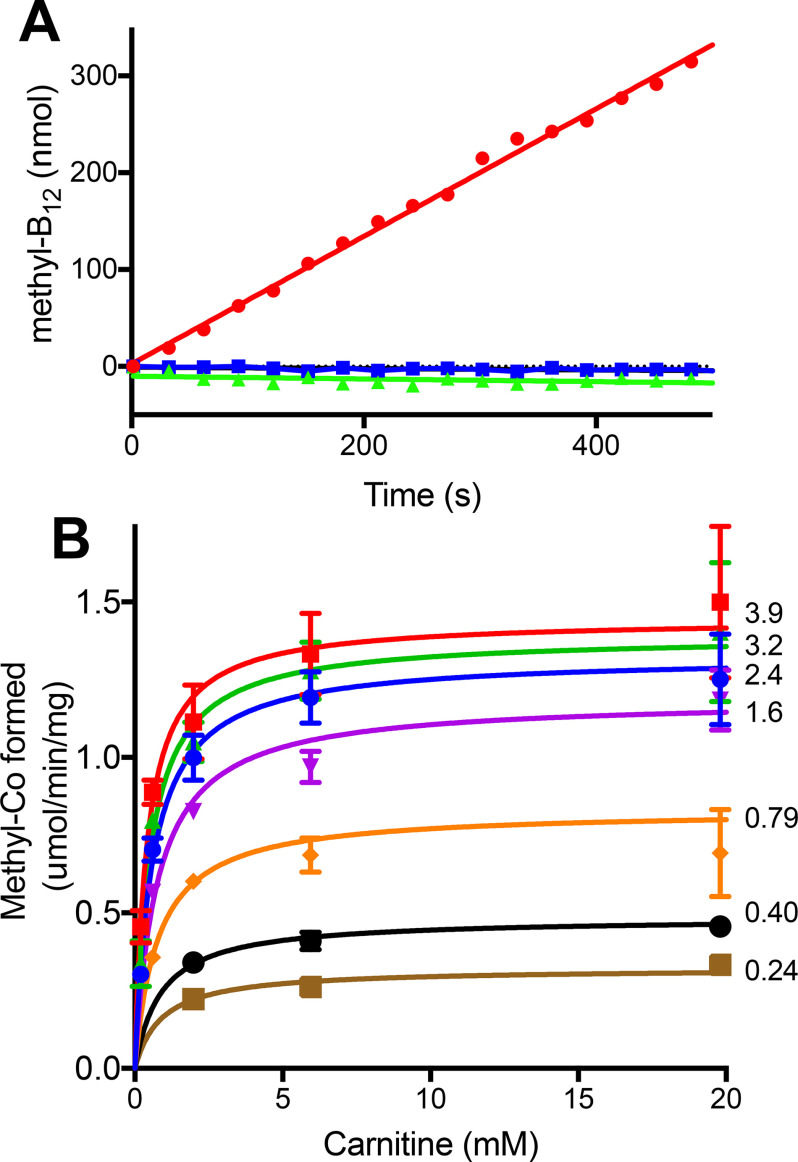
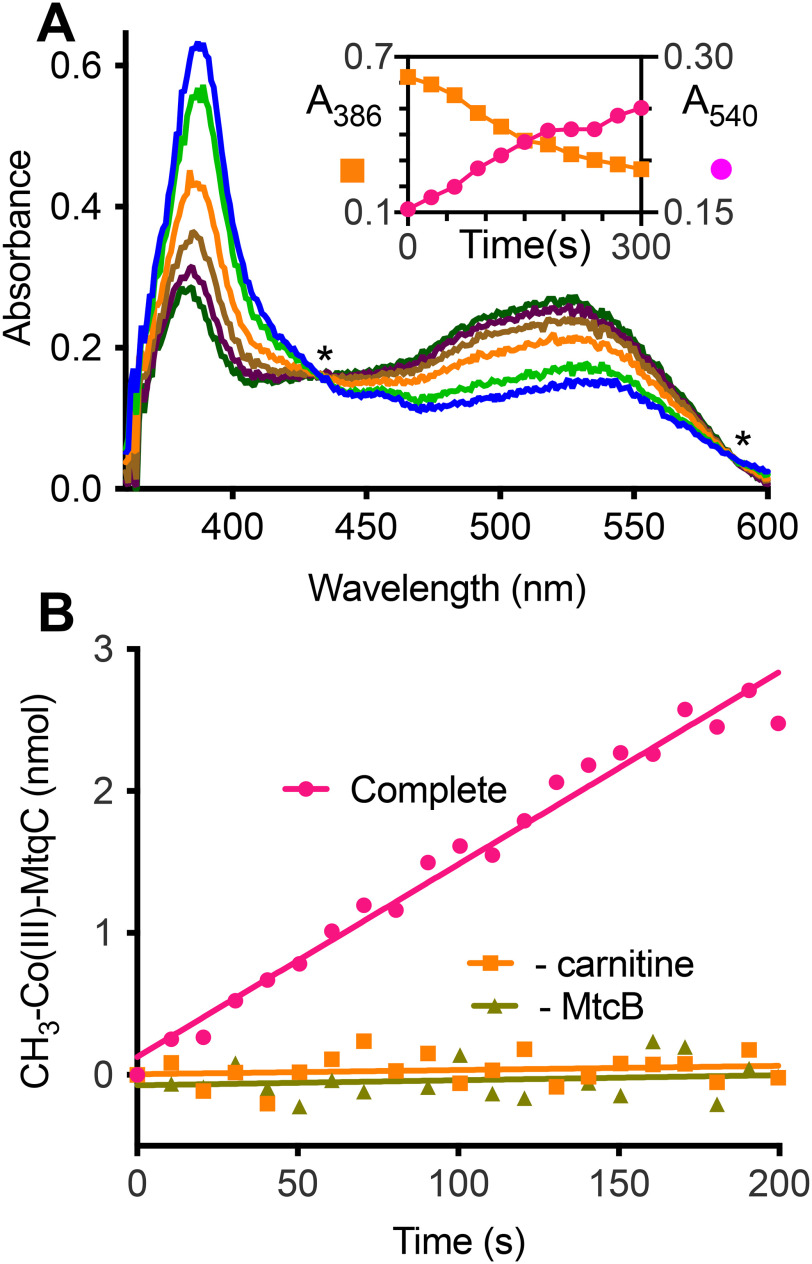
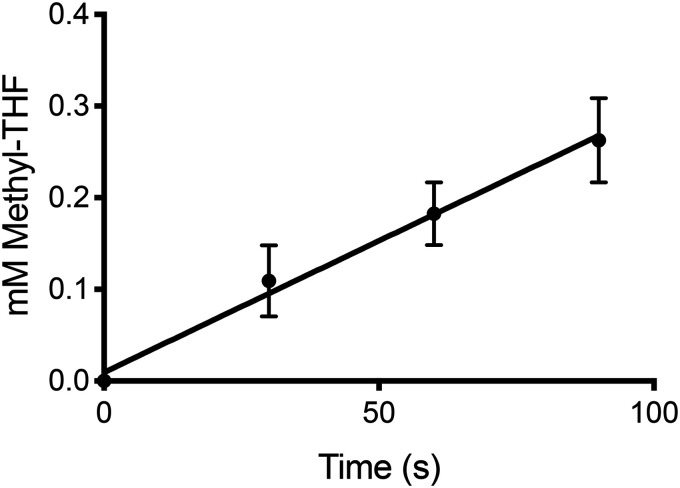
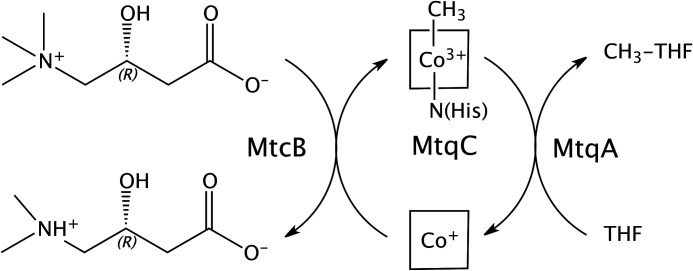
The trimethylamine methyltransferase MttB is the first described member of a superfamily comprising thousands of microbial proteins. Most members of the MttB superfamily are encoded by genes that lack the codon for pyrrolysine characteristic of trimethylamine methyltransferases, raising questions about the activities of these proteins. The superfamily member MtcB is found in the human intestinal isolate Eubacterium limosum ATCC 8486, an acetogen that can grow by demethylation of l-carnitine. Here, we demonstrate that MtcB catalyzes l-carnitine demethylation. When growing on l-carnitine, E. limosum excreted the unusual biological product norcarnitine as well as acetate, butyrate, and caproate. Cellular extracts of E. limosum grown on l-carnitine, but not lactate, methylated cob-(I)alamin or tetrahydrofolate using l-carnitine as methyl donor. MtcB, along with the corrinoid protein MtqC and the methylcorrinoid:tetrahydrofolate methyltransferase MtqA, were much more abundant in E. limosum cells grown on l-carnitine than on lactate. Recombinant MtcB methylates either cob(I)alamin or Co(I)-MtqC in the presence of l-carnitine and, to a much lesser extent, γ-butyrobetaine. Other quaternary amines were not substrates. Recombinant MtcB, MtqC, and MtqA methylated tetrahydrofolate via l-carnitine, forming a key intermediate in the acetogenic Wood-Ljungdahl pathway. To our knowledge, MtcB methylation of cobalamin or Co(I)-MtqC represents the first described mechanism of biological l-carnitine demethylation. The conversion of l-carnitine and its derivative γ-butyrobetaine to trimethylamine by the gut microbiome has been linked to cardiovascular disease. The activities of MtcB and related proteins in E. limosum might demethylate proatherogenic quaternary amines and contribute to the perceived health benefits of this human gut symbiont.
Keywords: acetogenesis; bacterial metabolism; carnitine; cobalamin; energy metabolism; enzyme catalysis; folate; microbiology; microbiome; one-carbon metabolism.
© 2020 Kountz et al.
Conflict of interest statement
Conflict of interest—The authors declare that they have no conflicts of interest with the contents of this article.
Figures
Comment in
-
Taking the "Me" out of meat: A new demethylation pathway dismantles a toxin's precursor.J Biol Chem. 2020 Aug 21;295(34):11982-11983. doi: 10.1074/jbc.H120.015332. J Biol Chem. 2020. PMID: 32826335 Free PMC article.
References
-
- Ferguson D. J. Jr., and Krzycki J. A. (1997) Reconstitution of trimethylamine-dependent coenzyme M methylation with the trimethylamine corrinoid protein and the isozymes of methyltransferase II from Methanosarcina barkeri. J. Bacteriol. 179, 846–852 10.1128/jb.179.3.846-852.1997 - DOI - PMC - PubMed
-
- Ticak T., Kountz D. J., Girosky K. E., Krzycki J. A., and Ferguson D. J. Jr. (2014) A nonpyrrolysine member of the widely distributed trimethylamine methyltransferase family is a glycine betaine methyltransferase. Proc. Natl. Acad. Sci. U. S. A. 111, E4668–E4676 10.1073/pnas.1409642111 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
