

A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis
- PMID: 32302292
- PMCID: PMC7253024
- DOI: 10.1172/jci.insight.132055
A human-origin probiotic cocktail ameliorates aging-related leaky gut and inflammation via modulating the microbiota/taurine/tight junction axis
Abstract
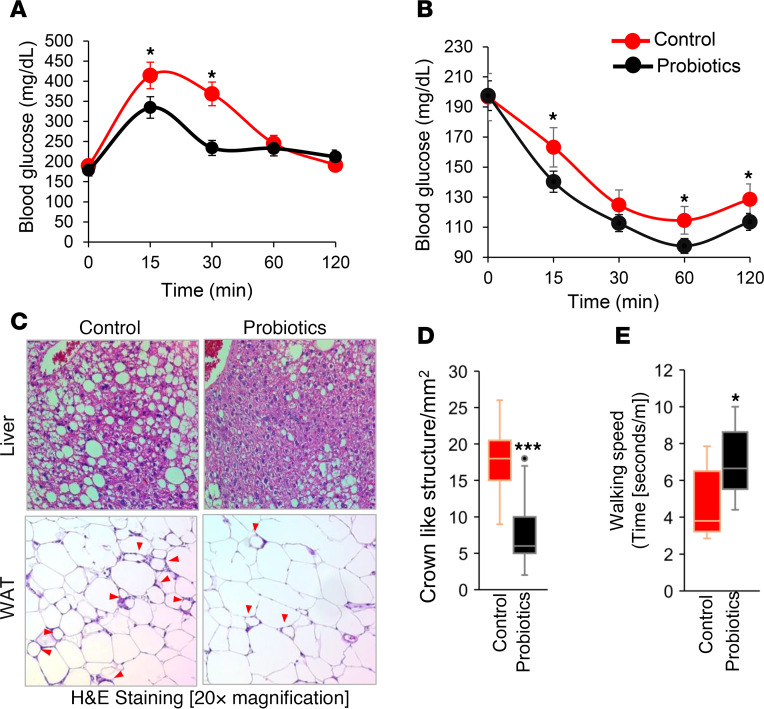
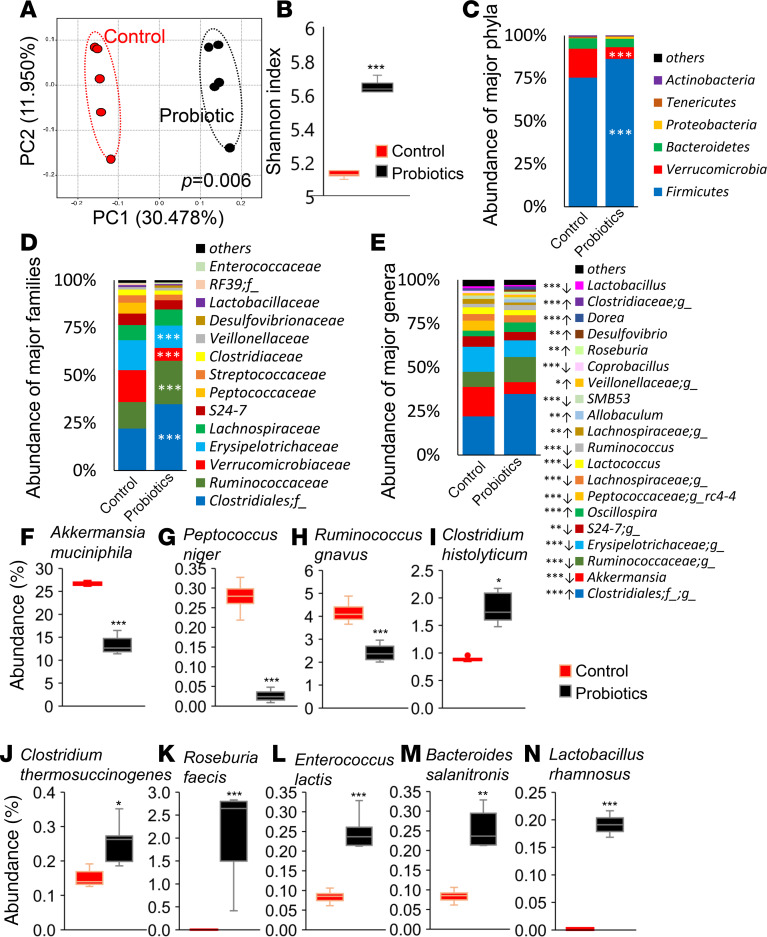
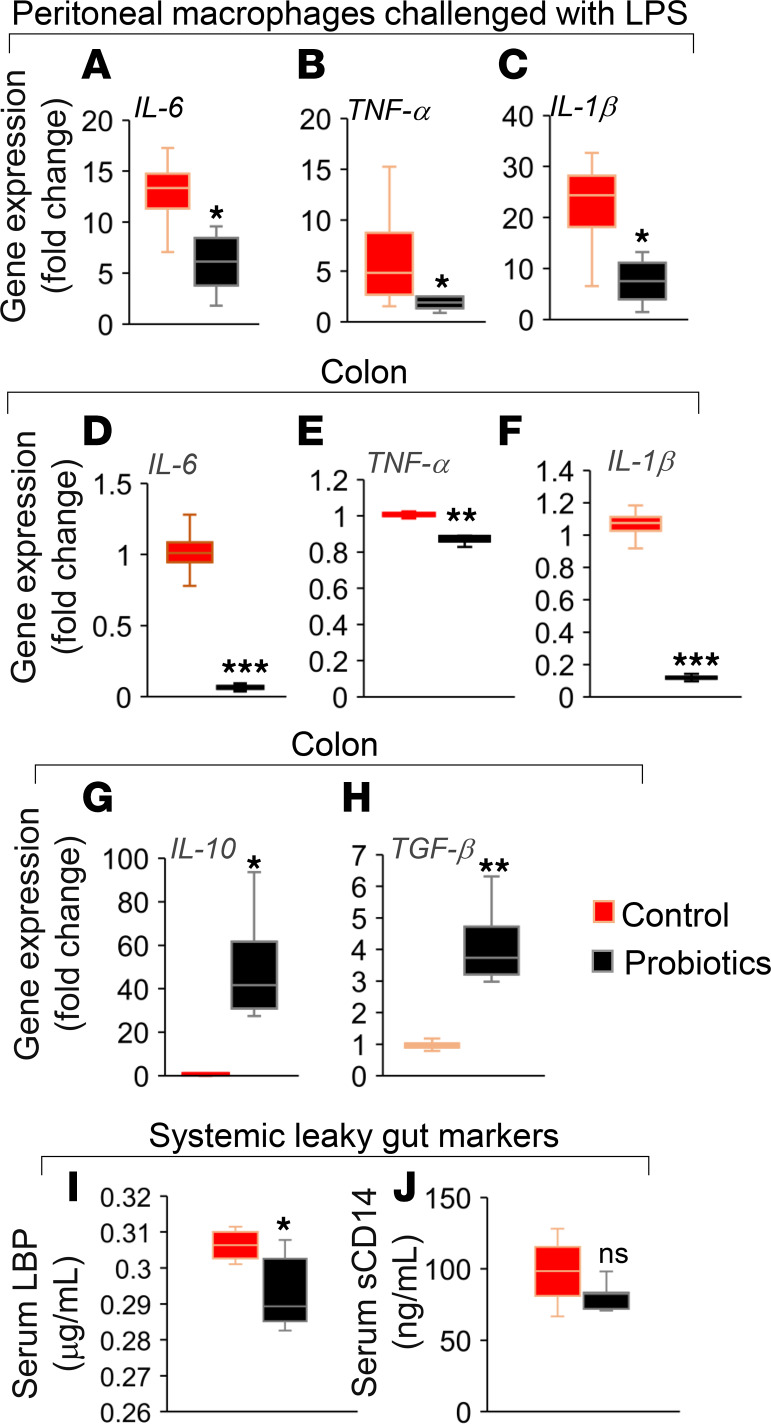
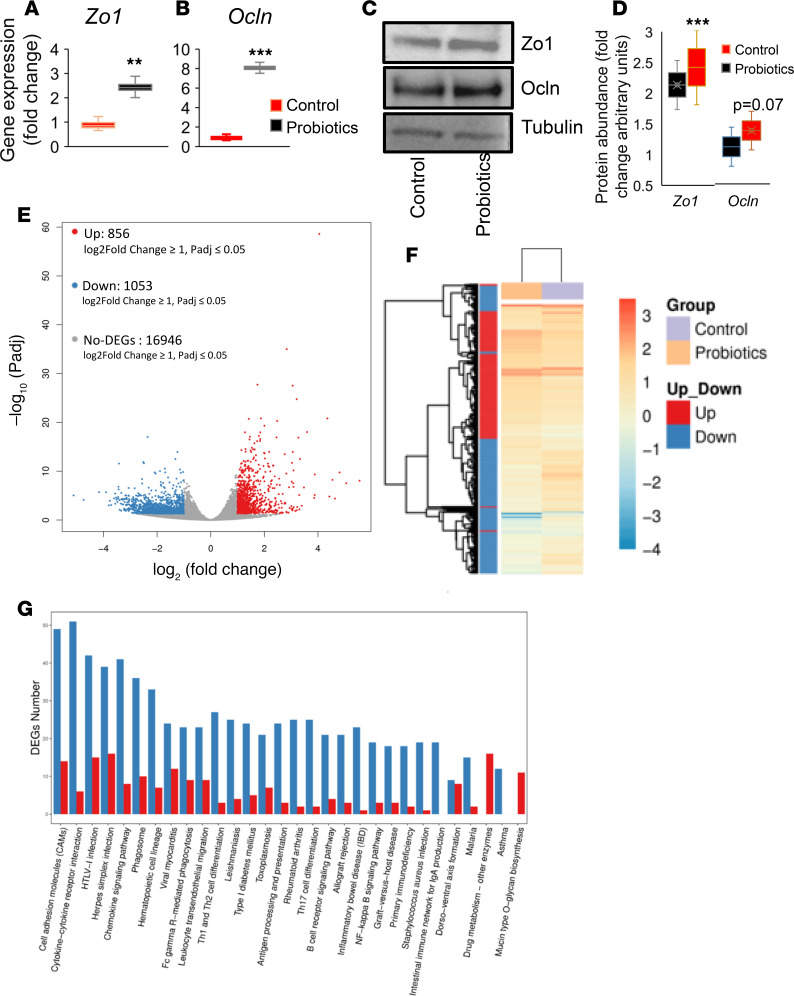
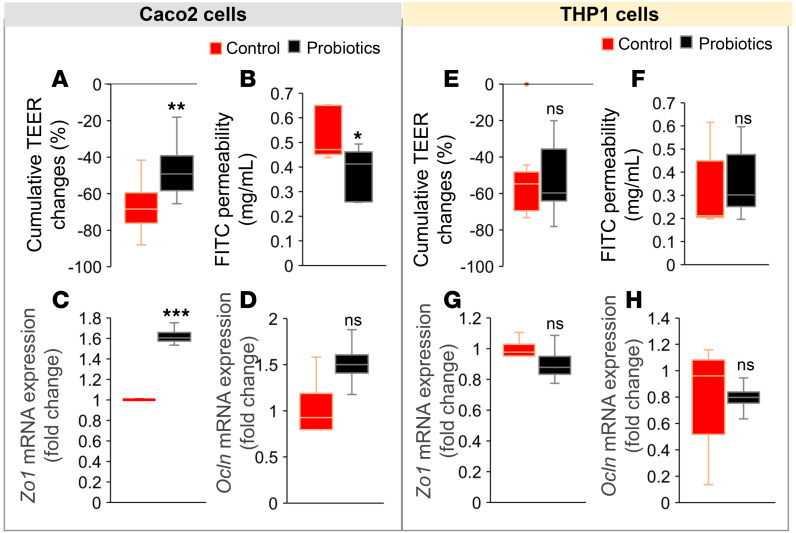
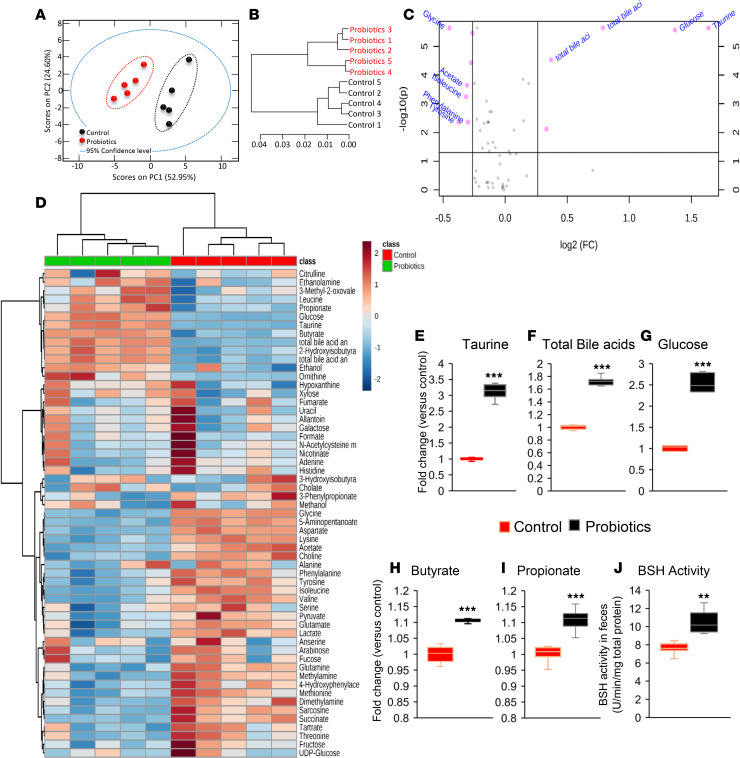
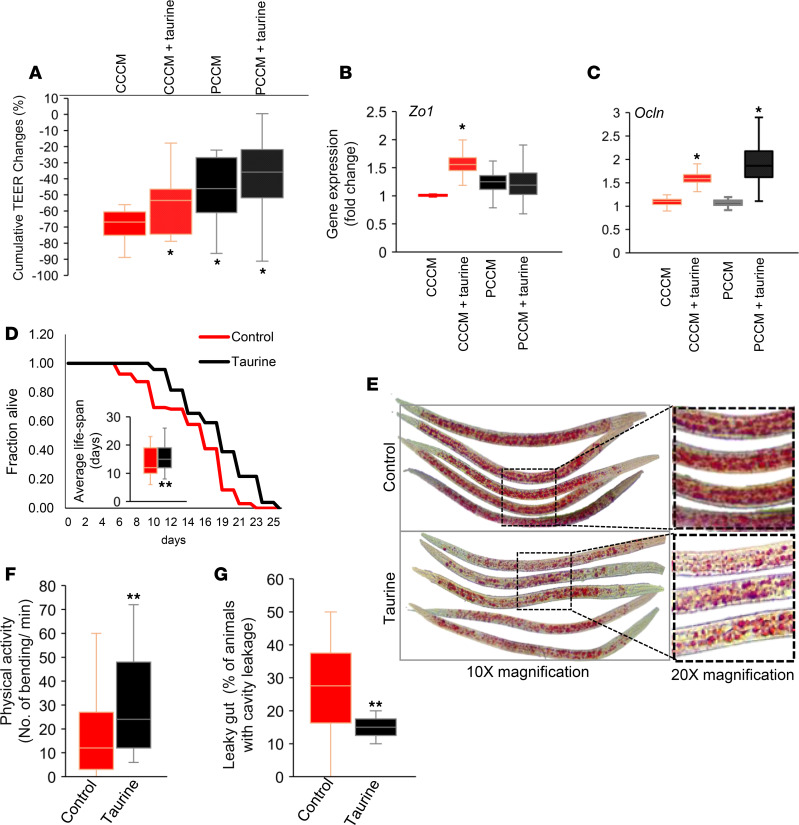
Inflammation is a major risk factor of morbidity and mortality in older adults. Although its precise etiology is unknown, low-grade inflammation in older adults is commonly associated with increased intestinal epithelial permeability (leaky gut) and abnormal (dysbiotic) gut microbiota. The increasing older population and lack of treatments to reduce aging-related microbiota dysbiosis, leaky gut, and inflammation culminates in a rise in aging-related comorbidities, constituting a significant public health concern. Here, we demonstrate that a human-origin probiotic cocktail containing 5 Lactobacillus and 5 Enterococcus strains isolated from healthy infant gut prevented high-fat diet-induced (HFD-induced) microbiota dysbiosis, leaky gut, inflammation, metabolic dysfunctions, and physical function decline in older mice. Probiotic-modulated gut microbiota primarily reduced leaky gut by increasing tight junctions, which in turn reduced inflammation. Mechanistically, probiotics modulated microbiota in a way to increase bile salt hydrolase activity, which in turn increased taurine abundance in the gut that stimulated tight junctions and suppressed gut leakiness. Furthermore, in Caenorhabditis elegans, taurine increased life span, reduced adiposity and leaky gut, and enhanced physical function. The results suggest that such probiotic therapies could prevent or treat aging-related leaky gut and inflammation in the elderly.
Keywords: Gastroenterology; Glucose metabolism; Innate immunity; Macrophages; Microbiology.
Conflict of interest statement
Figures
References
-
- Kavanagh K, Hsu FC, Davis AT, Kritchevsky SB, Rejeski WJ, Kim S. Biomarkers of leaky gut are related to inflammation and reduced physical function in older adults with cardiometabolic disease and mobility limitations. Geroscience. 2019;41(6):923–933. doi: 10.1007/s11357-019-00112-z. - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
