

Infection with genotoxin-producing Salmonella enterica synergises with loss of the tumour suppressor APC in promoting genomic instability via the PI3K pathway in colonic epithelial cells
- PMID: 31414579
- PMCID: PMC6899655
- DOI: 10.1111/cmi.13099
Infection with genotoxin-producing Salmonella enterica synergises with loss of the tumour suppressor APC in promoting genomic instability via the PI3K pathway in colonic epithelial cells
Abstract
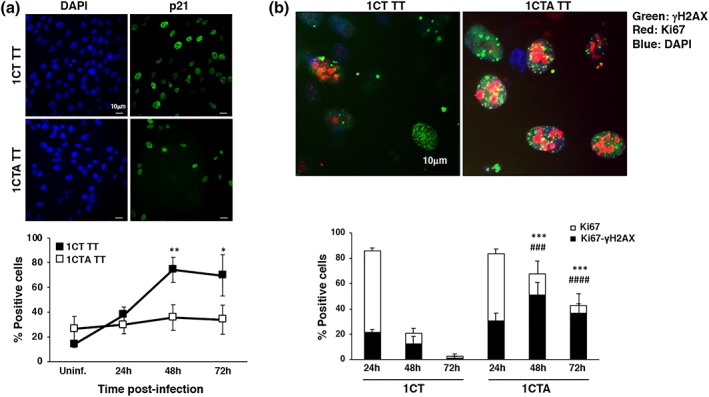
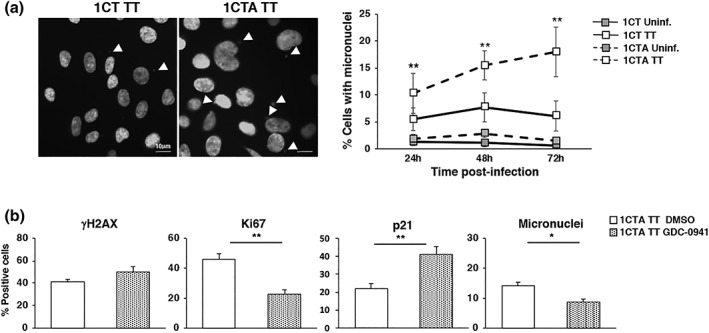
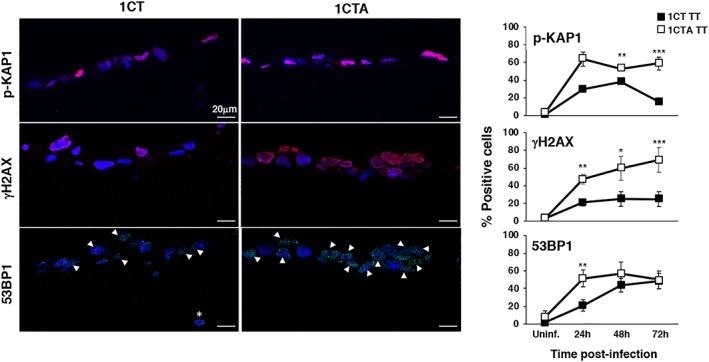
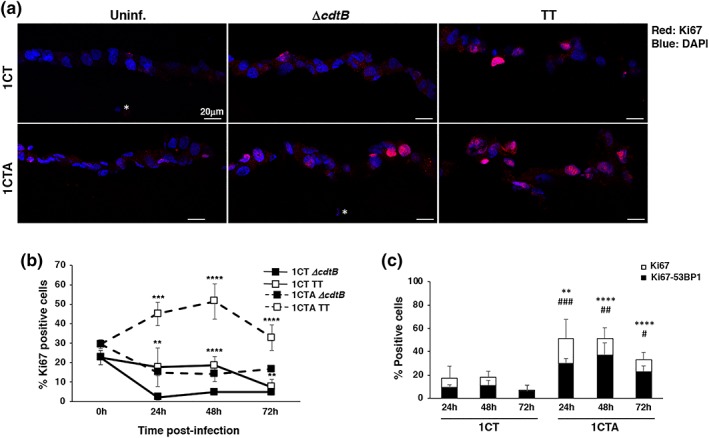
Several commensal and pathogenic Gram-negative bacteria produce DNA-damaging toxins that are considered bona fide carcinogenic agents. The microbiota of colorectal cancer (CRC) patients is enriched in genotoxin-producing bacteria, but their role in the pathogenesis of CRC is poorly understood. The adenomatous polyposis coli (APC) gene is mutated in familial adenomatous polyposis and in the majority of sporadic CRCs. We investigated whether the loss of APC alters the response of colonic epithelial cells to infection by Salmonella enterica, the only genotoxin-producing bacterium associated with cancer in humans. Using 2D and organotypic 3D cultures, we found that APC deficiency was associated with sustained activation of the DNA damage response, reduced capacity to repair different types of damage, including DNA breaks and oxidative damage, and failure to induce cell cycle arrest. The reduced DNA repair capacity and inability to activate adequate checkpoint responses was associated with increased genomic instability in APC-deficient cells exposed to the genotoxic bacterium. Inhibition of the checkpoint response was dependent on activation of the phosphatidylinositol 3-kinase pathway. These findings highlight the synergistic effect of the loss of APC and infection with genotoxin-producing bacteria in promoting a microenvironment conducive to malignant transformation.
Keywords: APC; DNA damage response; DNA repair; bacteria and cancer; bacterial genotoxin; organotypic model; tumour-suppressor gene.
© 2019 The Authors. Cellular Microbiology published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest
Figures
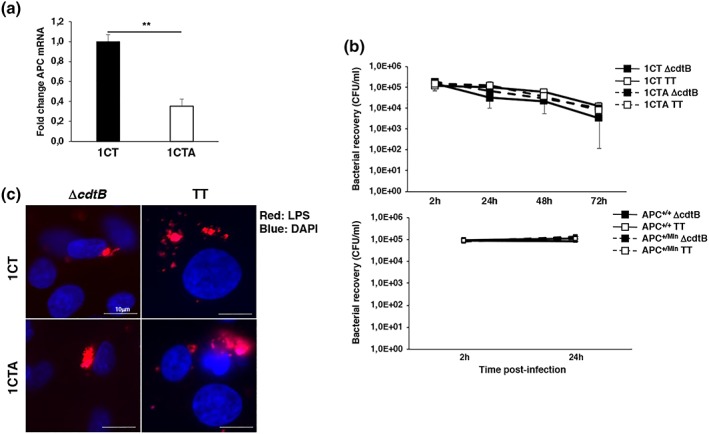
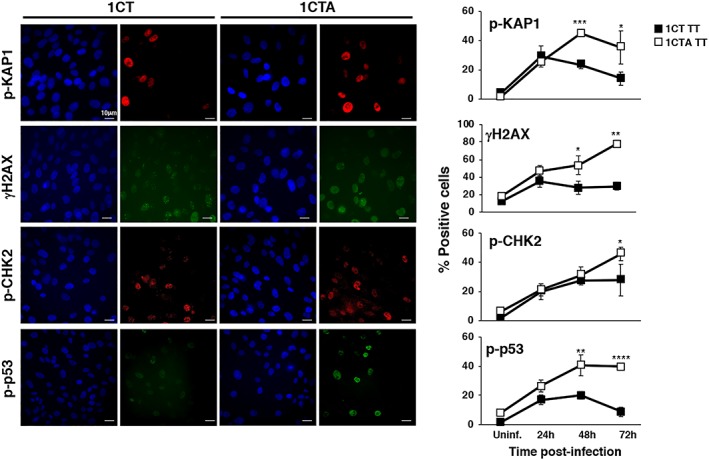
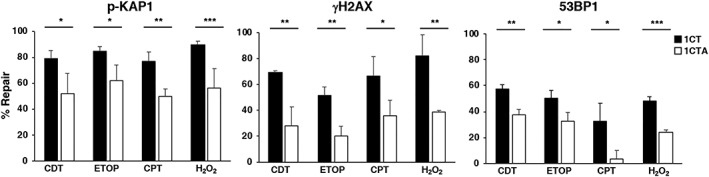
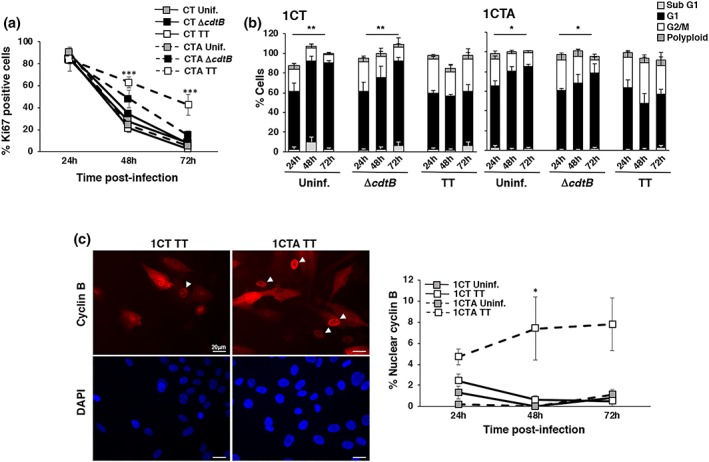
References
-
- Blazkova, H. , Krejcikova, K. , Moudry, P. , Frisan, T. , Hodny, Z. , & Bartek, J. (2010). Bacterial intoxication evokes cellular senescence with persistent DNA damage and cytokine signalling. Journal of Cellular and Molecular Medicine, 14(1‐2), 357–367. doi: JCMM862 [pii]. 10.1111/j.1582-4934.2009.00862.x - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
