

Microbiota derived short chain fatty acids promote histone crotonylation in the colon through histone deacetylases
- PMID: 29317660
- PMCID: PMC5760624
- DOI: 10.1038/s41467-017-02651-5
Microbiota derived short chain fatty acids promote histone crotonylation in the colon through histone deacetylases
Abstract
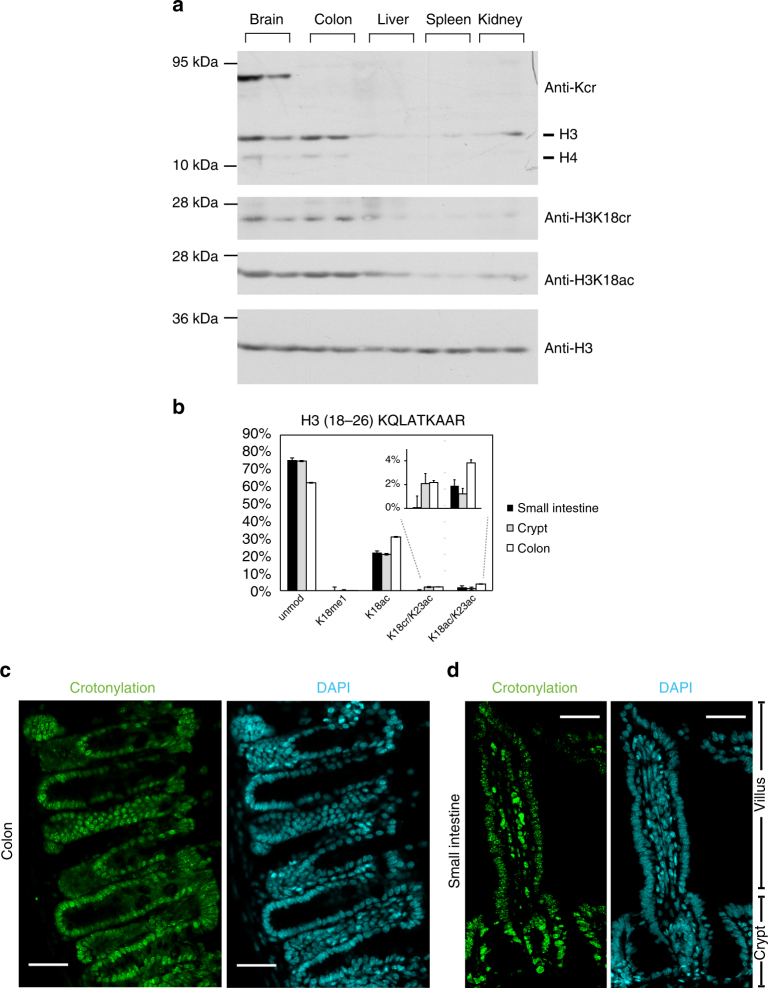
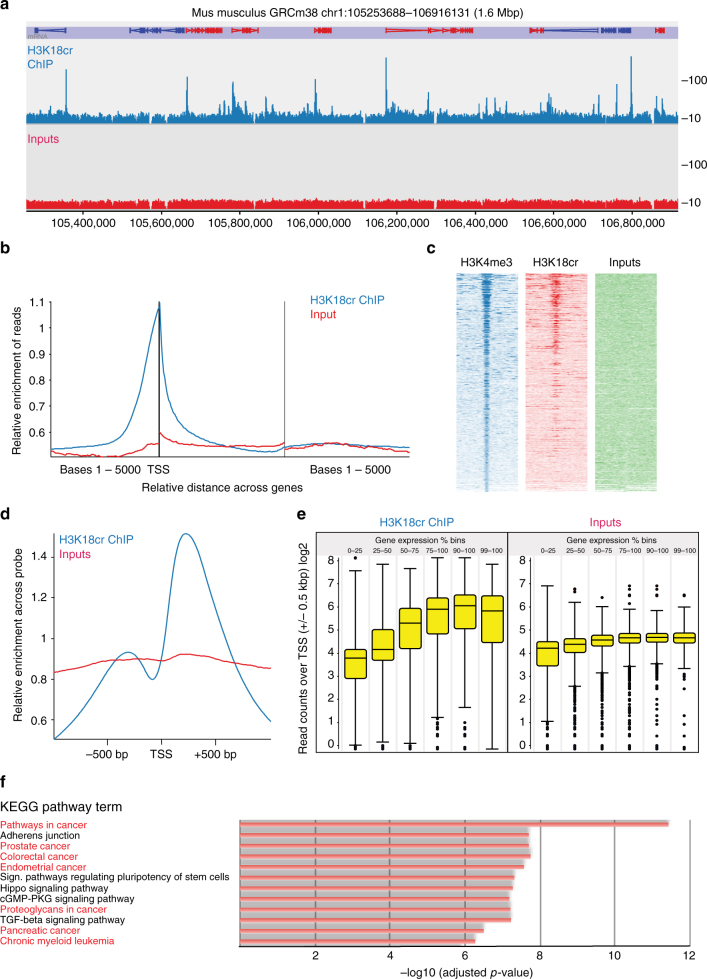
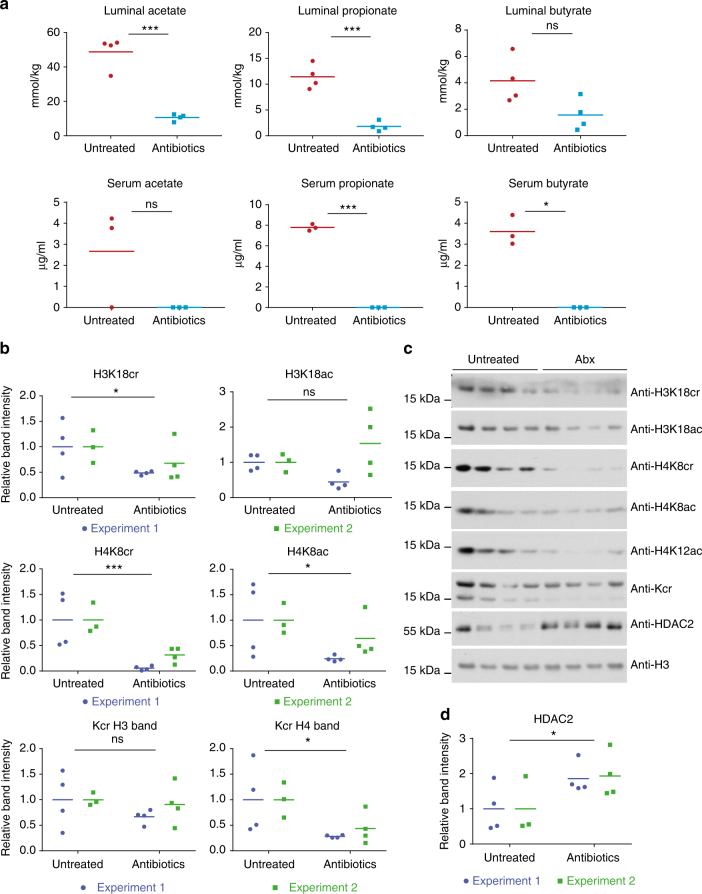
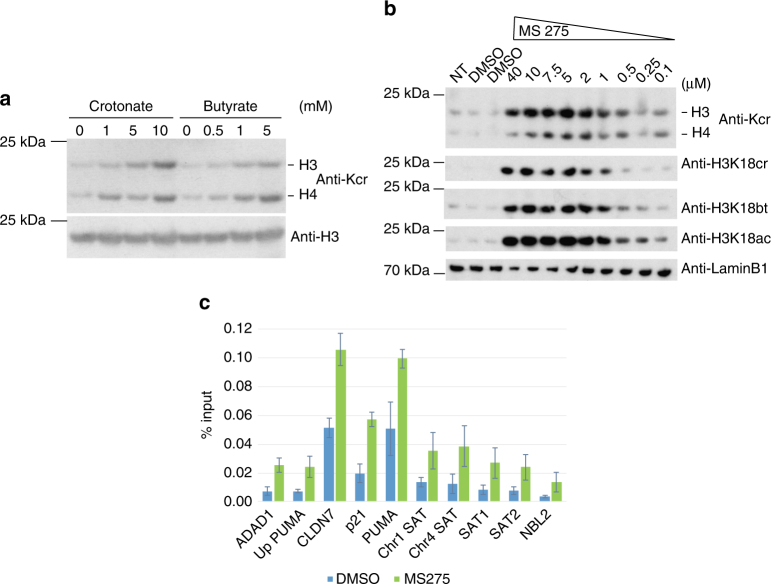
The recently discovered histone post-translational modification crotonylation connects cellular metabolism to gene regulation. Its regulation and tissue-specific functions are poorly understood. We characterize histone crotonylation in intestinal epithelia and find that histone H3 crotonylation at lysine 18 is a surprisingly abundant modification in the small intestine crypt and colon, and is linked to gene regulation. We show that this modification is highly dynamic and regulated during the cell cycle. We identify class I histone deacetylases, HDAC1, HDAC2, and HDAC3, as major executors of histone decrotonylation. We show that known HDAC inhibitors, including the gut microbiota-derived butyrate, affect histone decrotonylation. Consistent with this, we find that depletion of the gut microbiota leads to a global change in histone crotonylation in the colon. Our results suggest that histone crotonylation connects chromatin to the gut microbiota, at least in part, via short-chain fatty acids and HDACs.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
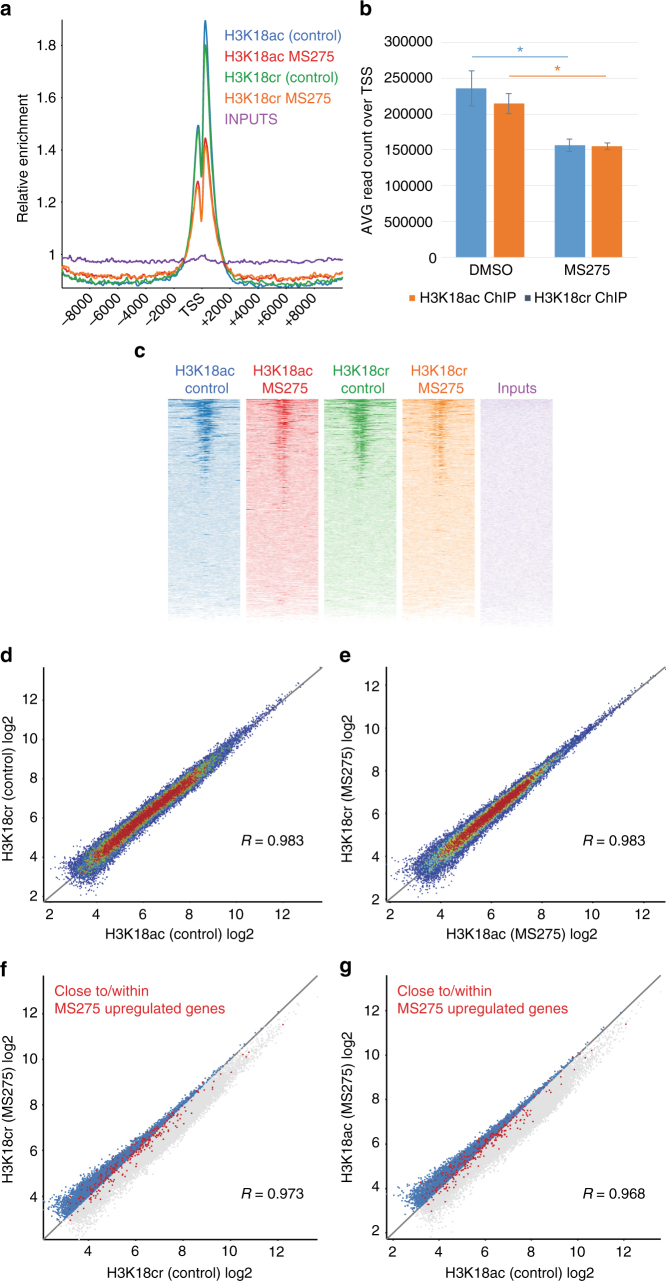
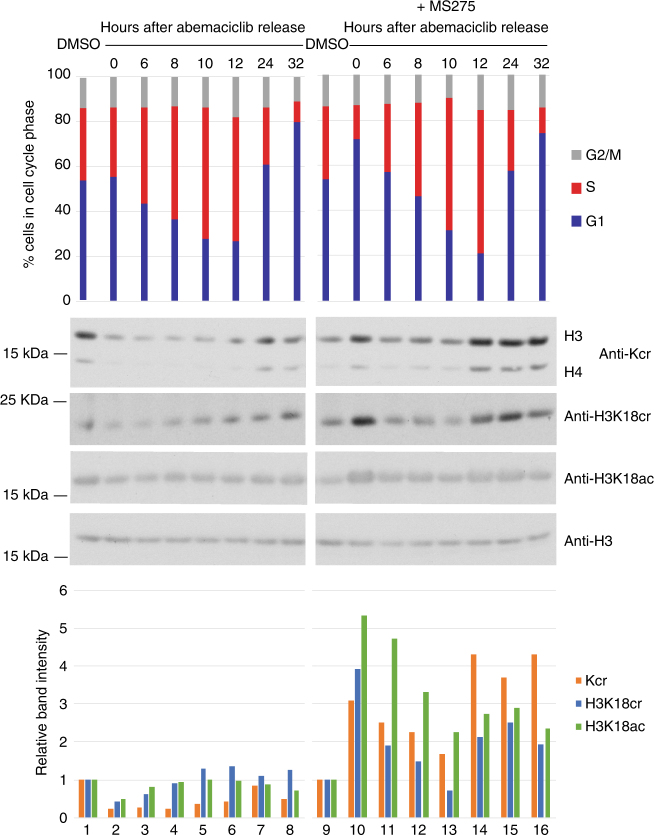
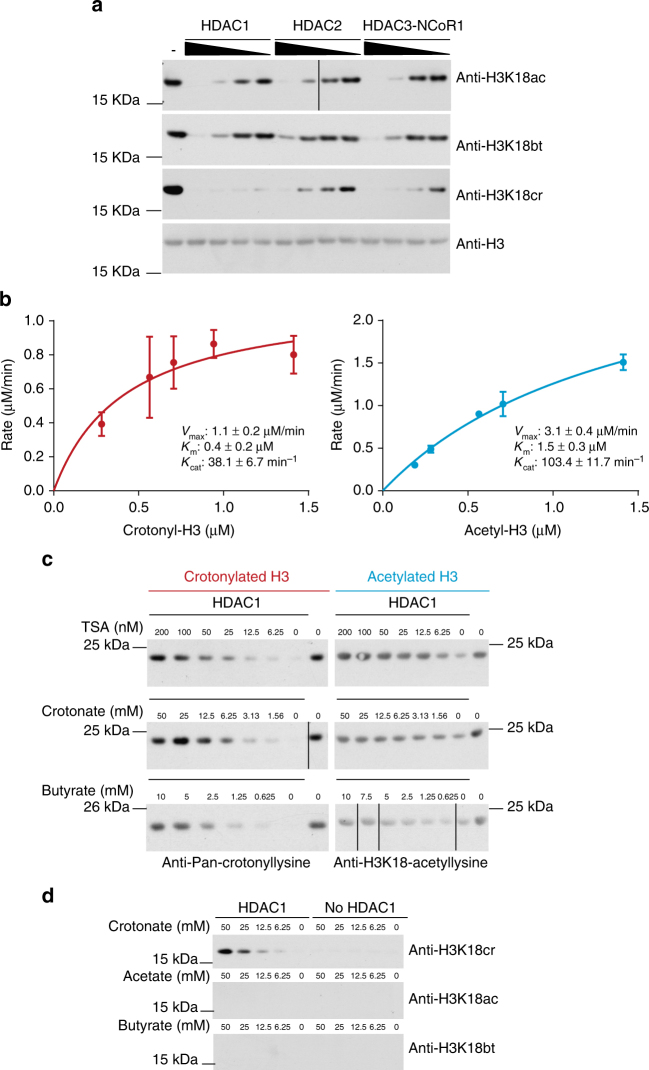
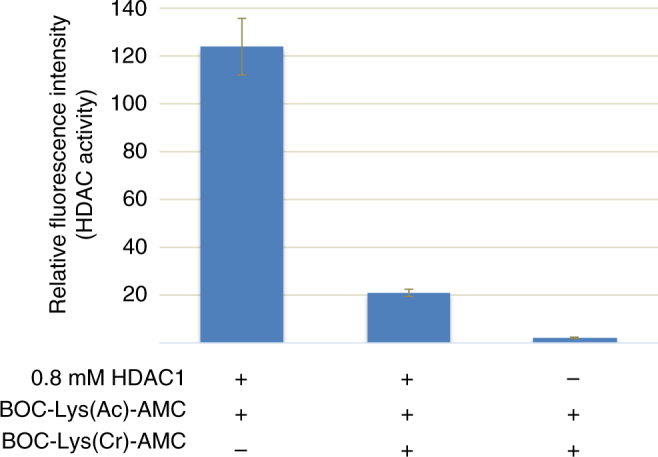
References
Publication types
MeSH terms
Substances
Grants and funding
- BBS/E/B/000C0409/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/F020236/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- NC/L001217/1/NC3RS_/National Centre for the Replacement, Refinement and Reduction of Animals in Research/United Kingdom
- MR/N009398/1/MRC_/Medical Research Council/United Kingdom
- G0701175/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
