

The Landscape of Type VI Secretion across Human Gut Microbiomes Reveals Its Role in Community Composition
- PMID: 28910638
- PMCID: PMC5679258
- DOI: 10.1016/j.chom.2017.08.010
The Landscape of Type VI Secretion across Human Gut Microbiomes Reveals Its Role in Community Composition
Abstract
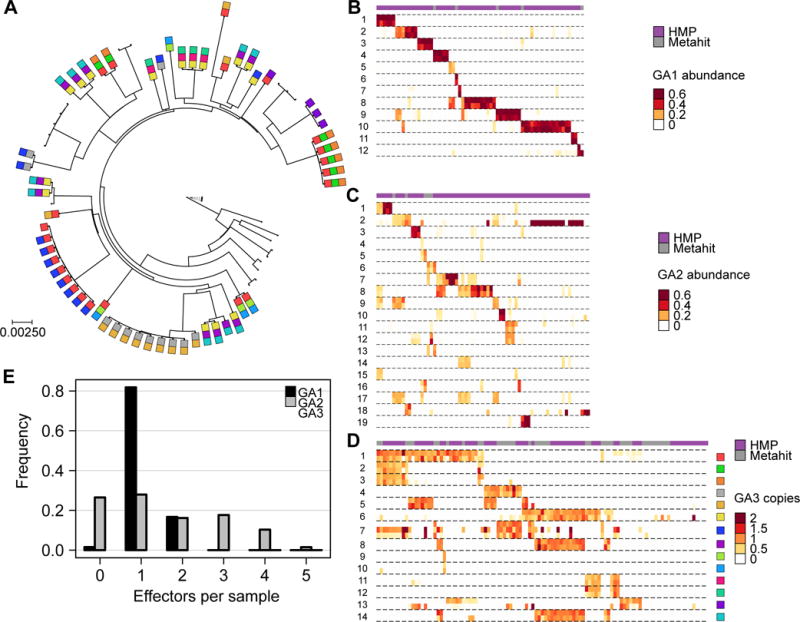
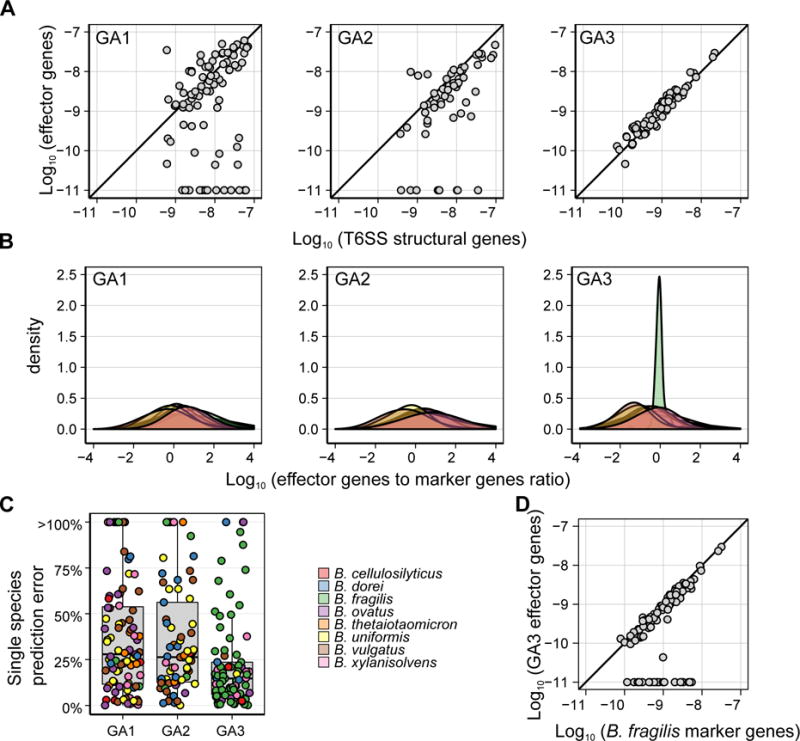
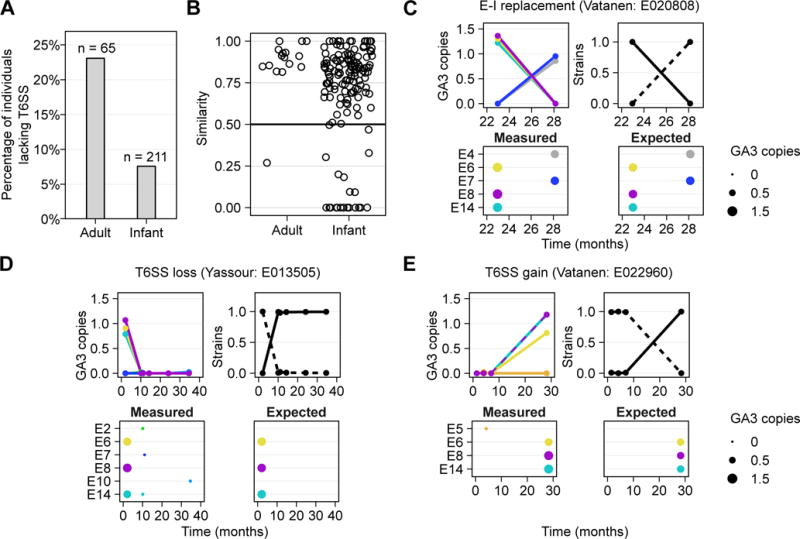
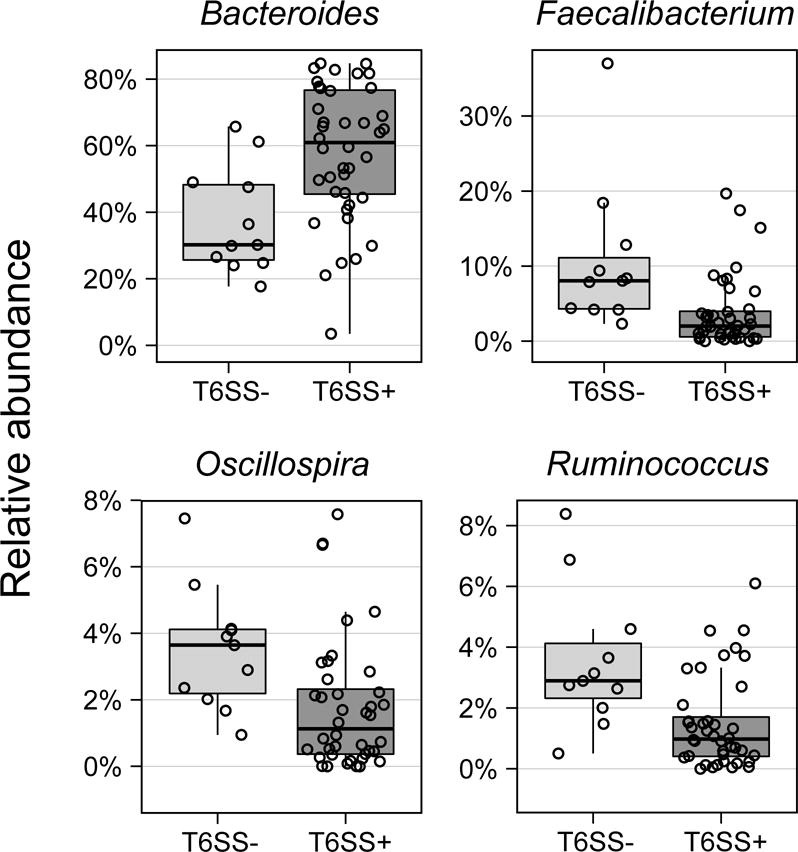
Although gut microbiome composition is well defined, the mechanisms underlying community assembly remain poorly understood. Bacteroidales possess three genetic architectures (GA1-3) of the type VI secretion system (T6SS), an effector delivery pathway that mediates interbacterial competition. Here we define the distribution and role of GA1-3 in the human gut using metagenomic analysis. We find that adult microbiomes harbor limited effector and cognate immunity genes, suggesting selection for compatibility at the species (GA1 and GA2) and strain (GA3) levels. Bacteroides fragilis GA3 is known to mediate potent inter-strain competition, and we observe GA3 enrichment among strains colonizing infant microbiomes, suggesting competition early in life. Additionally, GA3 is associated with increased Bacteroides abundance, indicating that this system confers an advantage in Bacteroides-rich ecosystems. Collectively, these analyses uncover the prevalence of T6SS-dependent competition and reveal its potential role in shaping human gut microbial composition.
Keywords: Bacteroidales; T6SS; human microbiome; infant microbiome; interbacterial competition; metagenomics; microbiome assembly; toxin; type VI secretion system.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Anderson MC, Vonaesch P, Saffarian A, Marteyn BS, Sansonetti PJ. Shigella sonnei Encodes a Functional T6SS Used for Interbacterial Competition and Niche Occupancy. Cell Host Microbe. 2017;21:769–776 e763. - PubMed
-
- Backhed F, Roswall J, Peng Y, Feng Q, Jia H, Kovatcheva-Datchary P, Li Y, Xia Y, Xie H, Zhong H, et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe. 2015;17:690–703. - PubMed
-
- Benz J, Meinhart A. Antibacterial effector/immunity systems: it’s just the tip of the iceberg. Curr Opin Microbiol. 2014;17:1–10. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
