

CX3CL1 is up-regulated in the rat hippocampus during memory-associated synaptic plasticity
- PMID: 25161610
- PMCID: PMC4130185
- DOI: 10.3389/fncel.2014.00233
CX3CL1 is up-regulated in the rat hippocampus during memory-associated synaptic plasticity
Abstract
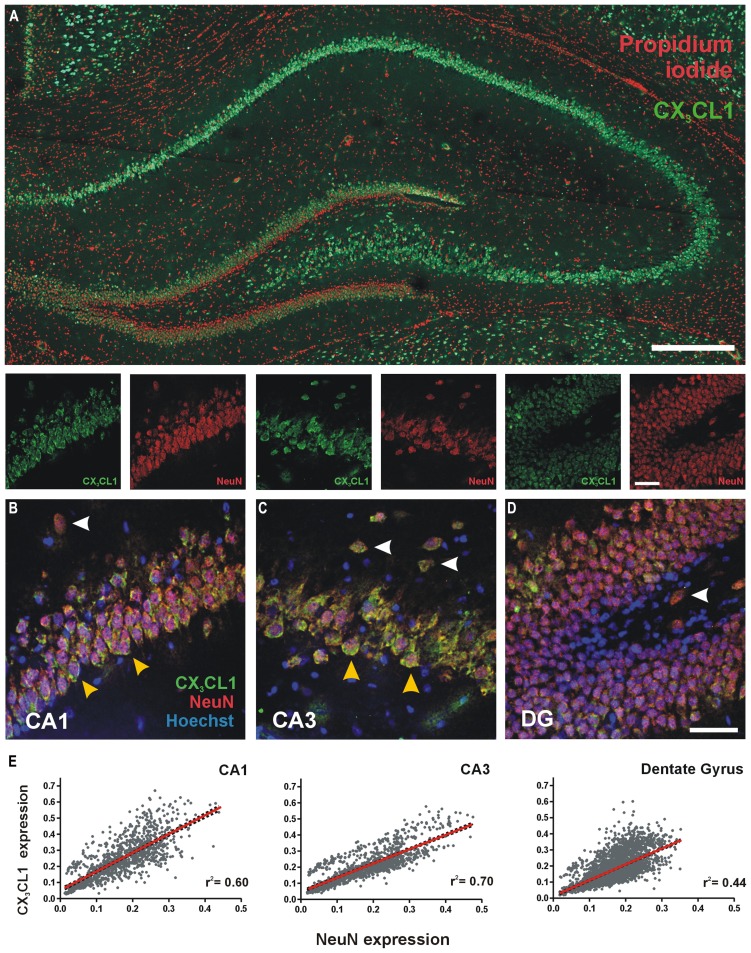
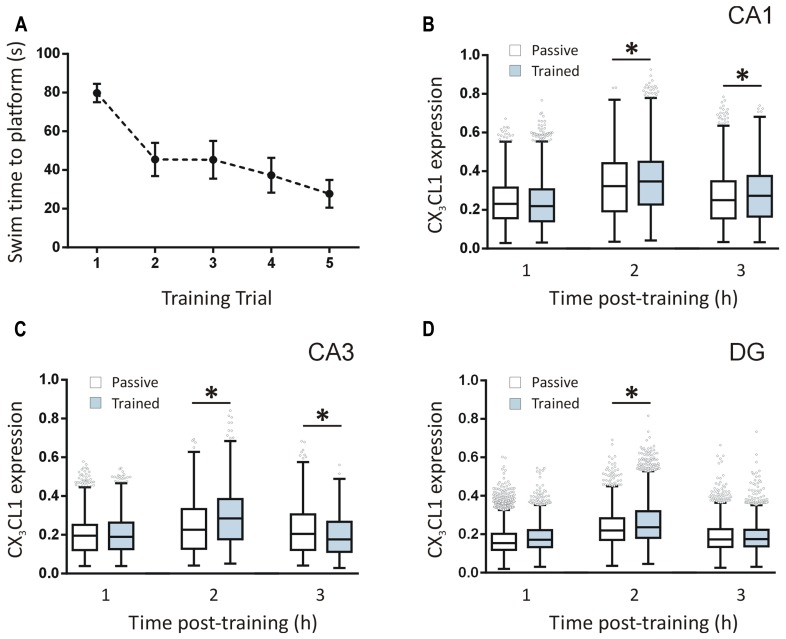
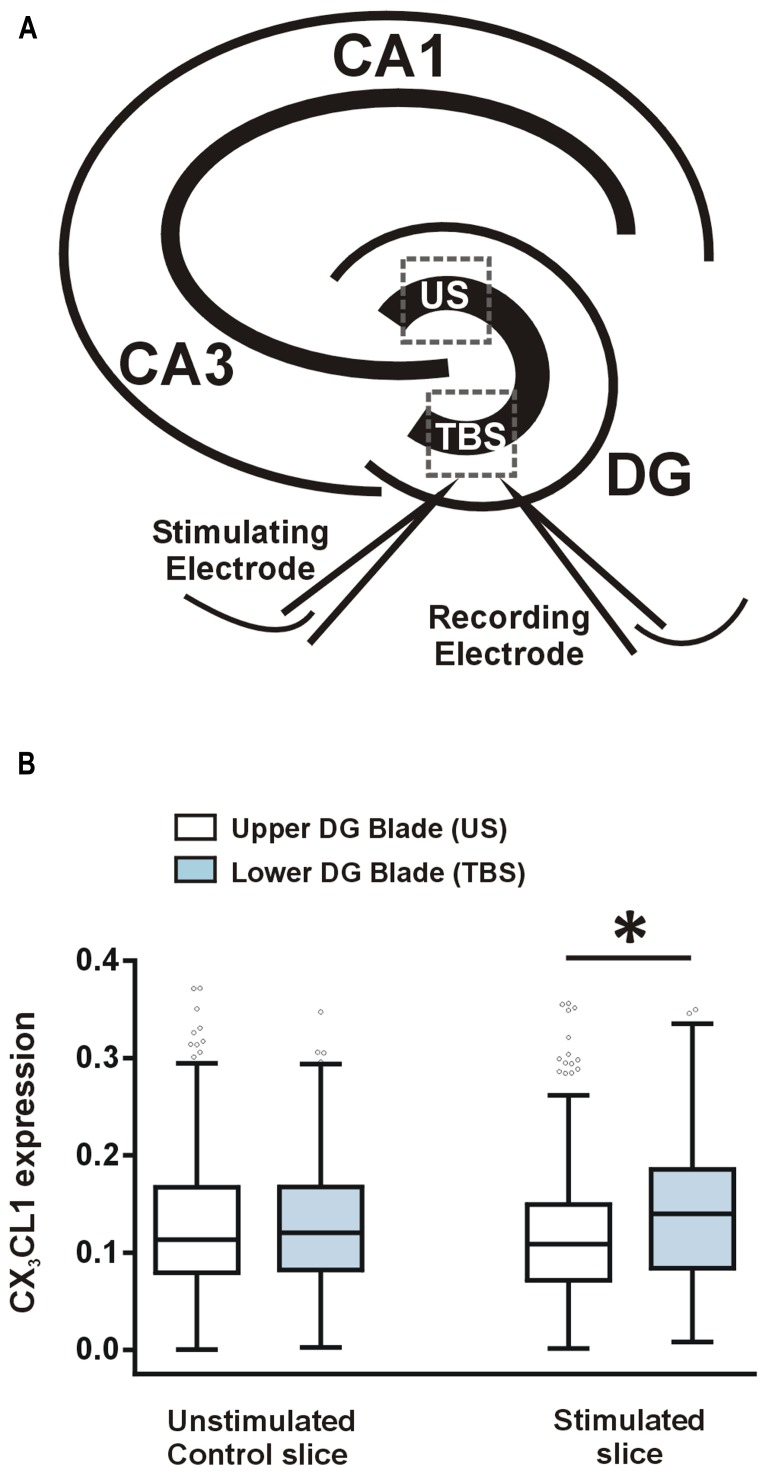
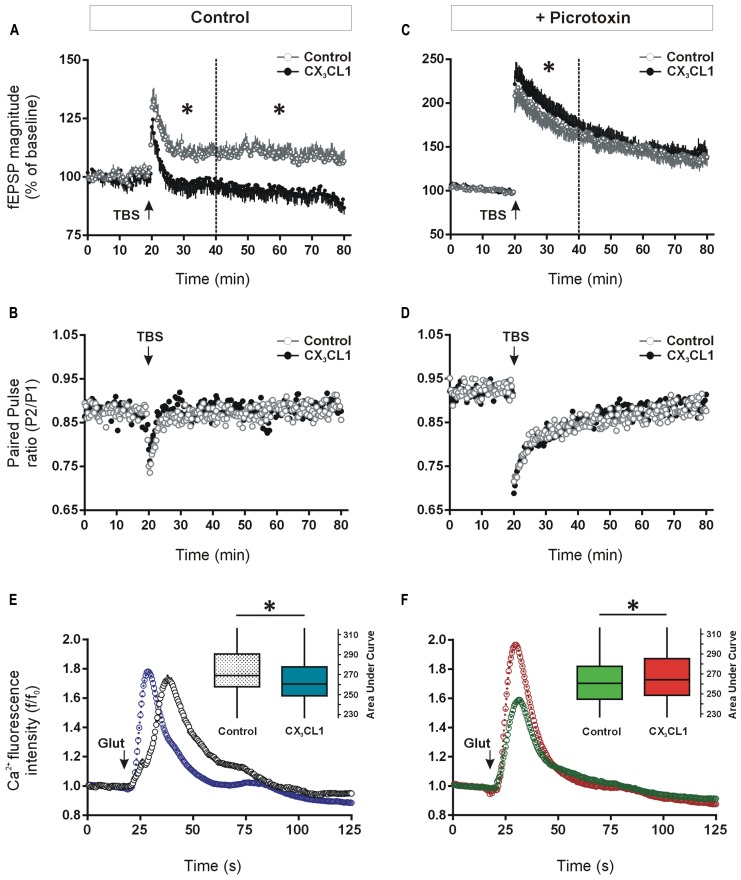
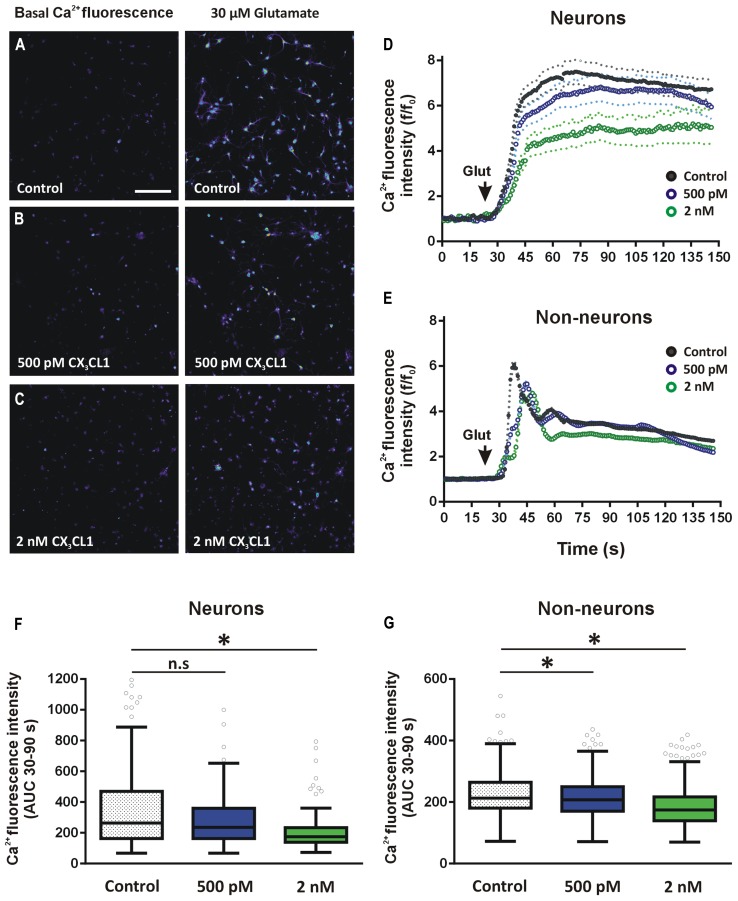
Several cytokines and chemokines are now known to play normal physiological roles in the brain where they act as key regulators of communication between neurons, glia, and microglia. In particular, cytokines and chemokines can affect cardinal cellular and molecular processes of hippocampal-dependent long-term memory consolidation including synaptic plasticity, synaptic scaling and neurogenesis. The chemokine, CX3CL1 (fractalkine), has been shown to modulate synaptic transmission and long-term potentiation (LTP) in the CA1 pyramidal cell layer of the hippocampus. Here, we confirm widespread expression of CX3CL1 on mature neurons in the adult rat hippocampus. We report an up-regulation in CX3CL1 protein expression in the CA1, CA3 and dentate gyrus (DG) of the rat hippocampus 2 h after spatial learning in the water maze task. Moreover, the same temporal increase in CX3CL1 was evident following LTP-inducing theta-burst stimulation in the DG. At physiologically relevant concentrations, CX3CL1 inhibited LTP maintenance in the DG. This attenuation in dentate LTP was lost in the presence of GABAA receptor/chloride channel antagonism. CX3CL1 also had opposing actions on glutamate-mediated rise in intracellular calcium in hippocampal organotypic slice cultures in the presence and absence of GABAA receptor/chloride channel blockade. Using primary dissociated hippocampal cultures, we established that CX3CL1 reduces glutamate-mediated intracellular calcium rises in both neurons and glia in a dose dependent manner. In conclusion, CX3CL1 is up-regulated in the hippocampus during a brief temporal window following spatial learning the purpose of which may be to regulate glutamate-mediated neurotransmission tone. Our data supports a possible role for this chemokine in the protective plasticity process of synaptic scaling.
Keywords: LTP; calcium imaging; chemokine signaling; fractalkine; learning and memory; water maze.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous