

Cardiorespiratory coupling: common rhythms in cardiac, sympathetic, and respiratory activities
- PMID: 24746049
- PMCID: PMC4052709
- DOI: 10.1016/B978-0-444-63274-6.00010-2
Cardiorespiratory coupling: common rhythms in cardiac, sympathetic, and respiratory activities
Abstract
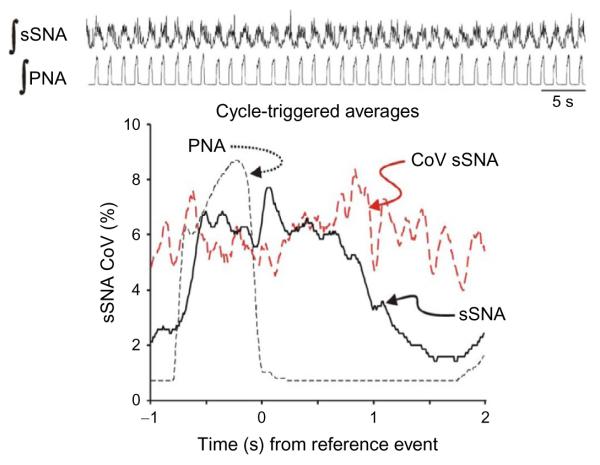
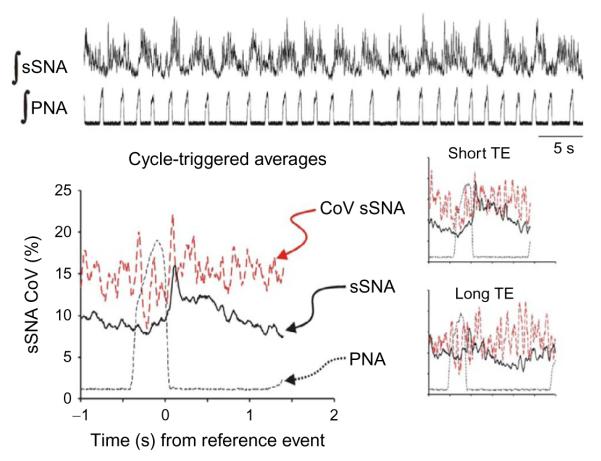
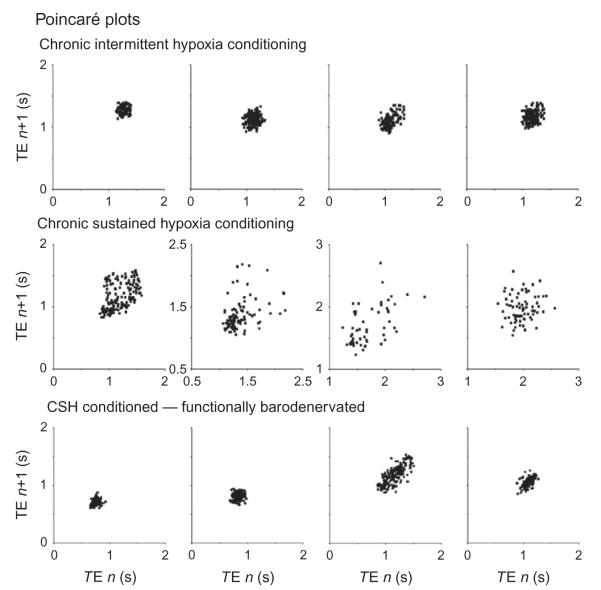
Cardiorespiratory coupling is an encompassing term describing more than the well-recognized influences of respiration on heart rate and blood pressure. Our data indicate that cardiorespiratory coupling reflects a reciprocal interaction between autonomic and respiratory control systems, and the cardiovascular system modulates the ventilatory pattern as well. For example, cardioventilatory coupling refers to the influence of heart beats and arterial pulse pressure on respiration and is the tendency for the next inspiration to start at a preferred latency after the last heart beat in expiration. Multiple complementary, well-described mechanisms mediate respiration's influence on cardiovascular function, whereas mechanisms mediating the cardiovascular system's influence on respiration may only be through the baroreceptors but are just being identified. Our review will describe a differential effect of conditioning rats with either chronic intermittent or sustained hypoxia on sympathetic nerve activity but also on ventilatory pattern variability. Both intermittent and sustained hypoxia increase sympathetic nerve activity after 2 weeks but affect sympatho-respiratory coupling differentially. Intermittent hypoxia enhances sympatho-respiratory coupling, which is associated with low variability in the ventilatory pattern. In contrast, after constant hypobaric hypoxia, 1-to-1 coupling between bursts of sympathetic and phrenic nerve activity is replaced by 2-to-3 coupling. This change in coupling pattern is associated with increased variability of the ventilatory pattern. After baro-denervating hypobaric hypoxic-conditioned rats, splanchnic sympathetic nerve activity becomes tonic (distinct bursts are absent) with decreases during phrenic nerve bursts and ventilatory pattern becomes regular. Thus, conditioning rats to either intermittent or sustained hypoxia accentuates the reciprocal nature of cardiorespiratory coupling. Finally, identifying a compelling physiologic purpose for cardiorespiratory coupling is the biggest barrier for recognizing its significance. Cardiorespiratory coupling has only a small effect on the efficiency of gas exchange; rather, we propose that cardiorespiratory control system may act as weakly coupled oscillator to maintain rhythms within a bounded variability.
Keywords: neural control of heart rate; neural control of respiration; neural control of sympathetic nerve activity; weakly coupled oscillators.
© 2014 Elsevier B.V. All rights reserved.
Figures

References
-
- Heart rate variability. Standards of measurement, physiological interpretation, and clinical use. Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Eur. Heart. J. 1996;17:354–381. - PubMed
-
- Baekey DM, Dick TE, Paton JF. Pontomedullary transection attenuates central respiratory modulation of sympathetic discharge, heart rate and the baroreceptor reflex in the in situ rat preparation. Exp. Physiol. 2008;93:803–816. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
