

Disruption of the Circadian Clock in Mice Increases Intestinal Permeability and Promotes Alcohol-Induced Hepatic Pathology and Inflammation
- PMID: 23825629
- PMCID: PMC3688973
- DOI: 10.1371/journal.pone.0067102
Disruption of the Circadian Clock in Mice Increases Intestinal Permeability and Promotes Alcohol-Induced Hepatic Pathology and Inflammation
Abstract
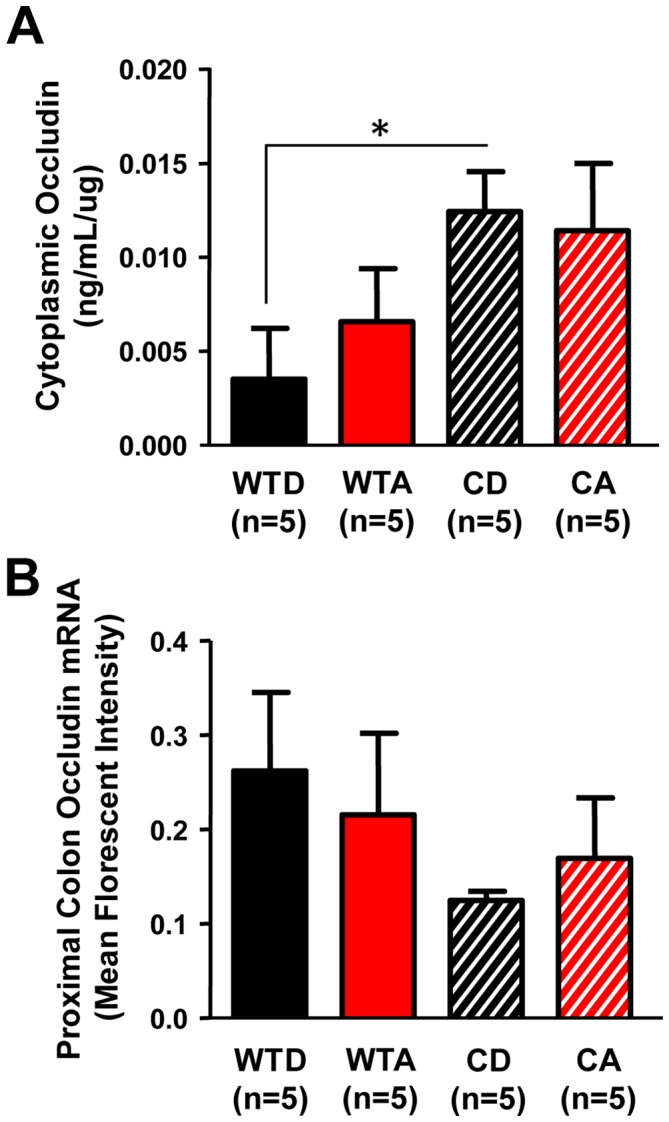
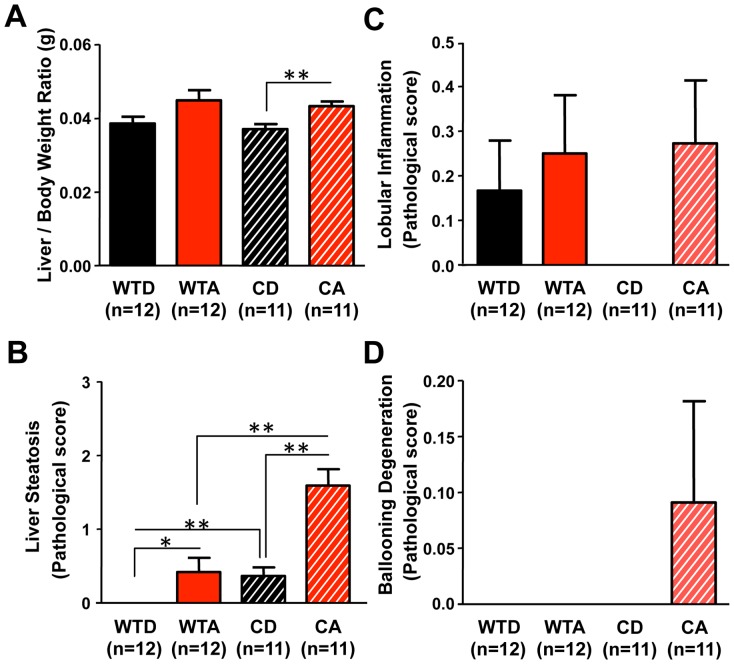
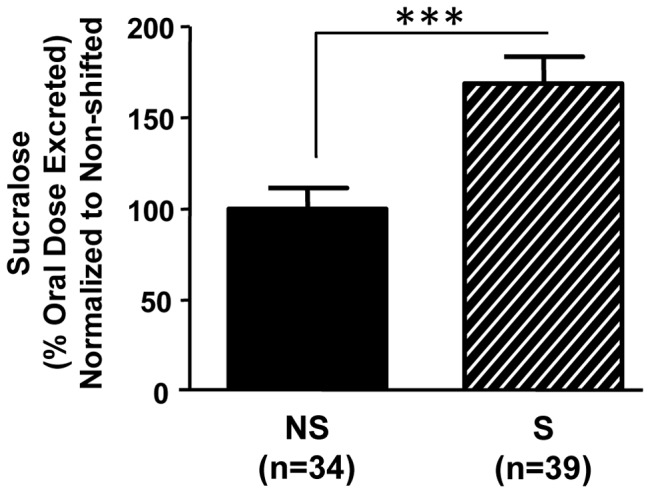
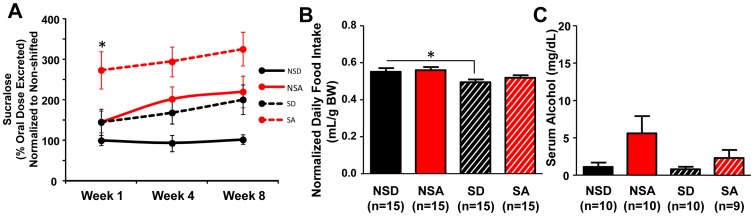
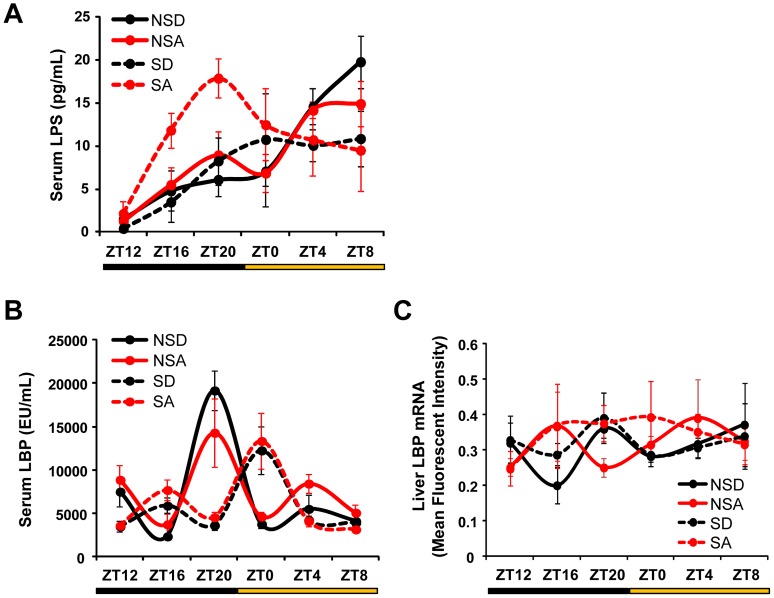
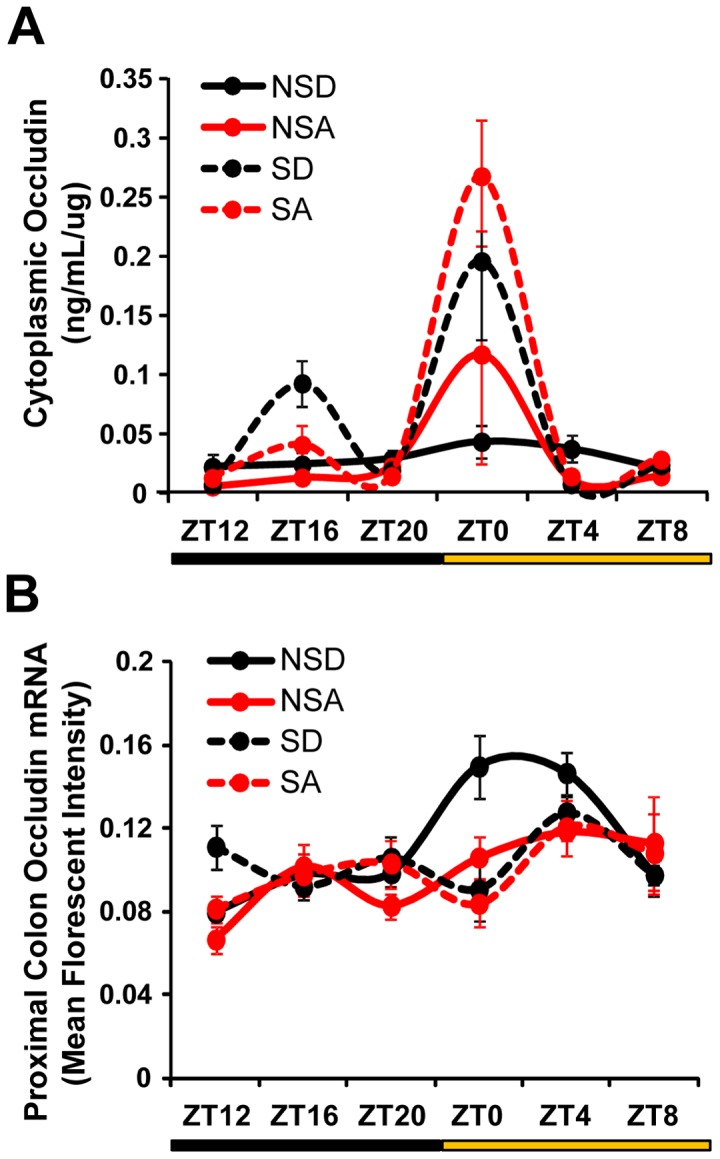
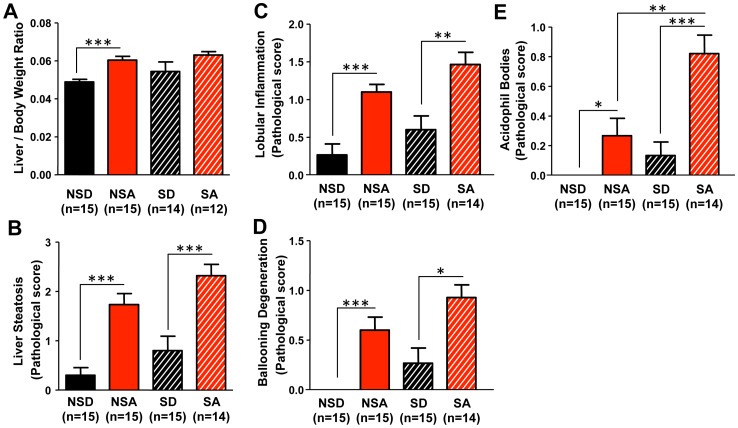
The circadian clock orchestrates temporal patterns of physiology and behavior relative to the environmental light:dark cycle by generating and organizing transcriptional and biochemical rhythms in cells and tissues throughout the body. Circadian clock genes have been shown to regulate the physiology and function of the gastrointestinal tract. Disruption of the intestinal epithelial barrier enables the translocation of proinflammatory bacterial products, such as endotoxin, across the intestinal wall and into systemic circulation; a process that has been linked to pathologic inflammatory states associated with metabolic, hepatic, cardiovascular and neurodegenerative diseases - many of which are commonly reported in shift workers. Here we report, for the first time, that circadian disorganization, using independent genetic and environmental strategies, increases permeability of the intestinal epithelial barrier (i.e., gut leakiness) in mice. Utilizing chronic alcohol consumption as a well-established model of induced intestinal hyperpermeability, we also found that both genetic and environmental circadian disruption promote alcohol-induced gut leakiness, endotoxemia and steatohepatitis, possibly through a mechanism involving the tight junction protein occludin. Circadian organization thus appears critical for the maintenance of intestinal barrier integrity, especially in the context of injurious agents, such as alcohol. Circadian disruption may therefore represent a previously unrecognized risk factor underlying the susceptibility to or development of alcoholic liver disease, as well as other conditions associated with intestinal hyperpermeability and an endotoxin-triggered inflammatory state.
Conflict of interest statement
Figures
References
-
- Hoogerwerf WA, Hellmich HL, Cornelissen G, Halberg F, Shahinian VB, et al. (2007) Clock gene expression in the murine gastrointestinal tract: endogenous rhythmicity and effects of a feeding regimen. Gastroenterology 133: 1250–1260. - PubMed
-
- Marra G, Anti M, Percesepe A, Armelao F, Ficarelli R, et al. (1994) Circadian variations of epithelial cell proliferation in human rectal crypts. Gastroenterology 106: 982–987. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
