

Central chemoreceptors: locations and functions
- PMID: 23728974
- PMCID: PMC4802370
- DOI: 10.1002/cphy.c100083
Central chemoreceptors: locations and functions
Abstract
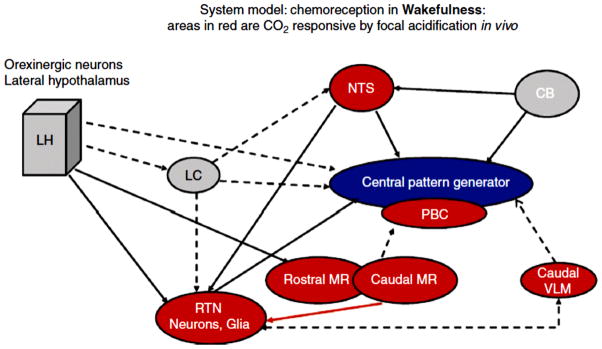
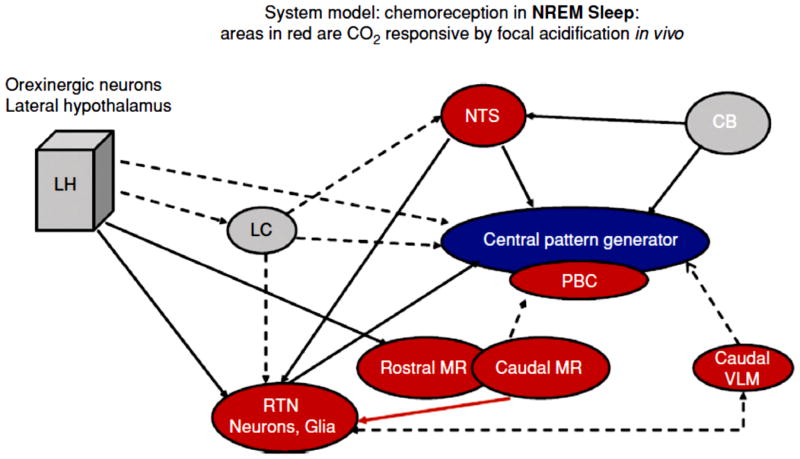
Central chemoreception traditionally refers to a change in ventilation attributable to changes in CO2/H(+) detected within the brain. Interest in central chemoreception has grown substantially since the previous Handbook of Physiology published in 1986. Initially, central chemoreception was localized to areas on the ventral medullary surface, a hypothesis complemented by the recent identification of neurons with specific phenotypes near one of these areas as putative chemoreceptor cells. However, there is substantial evidence that many sites participate in central chemoreception some located at a distance from the ventral medulla. Functionally, central chemoreception, via the sensing of brain interstitial fluid H(+), serves to detect and integrate information on (i) alveolar ventilation (arterial PCO2), (ii) brain blood flow and metabolism, and (iii) acid-base balance, and, in response, can affect breathing, airway resistance, blood pressure (sympathetic tone), and arousal. In addition, central chemoreception provides a tonic "drive" (source of excitation) at the normal, baseline PCO2 level that maintains a degree of functional connectivity among brainstem respiratory neurons necessary to produce eupneic breathing. Central chemoreception responds to small variations in PCO2 to regulate normal gas exchange and to large changes in PCO2 to minimize acid-base changes. Central chemoreceptor sites vary in function with sex and with development. From an evolutionary perspective, central chemoreception grew out of the demands posed by air versus water breathing, homeothermy, sleep, optimization of the work of breathing with the "ideal" arterial PCO2, and the maintenance of the appropriate pH at 37°C for optimal protein structure and function.
© 2012 American Physiological Society
Figures
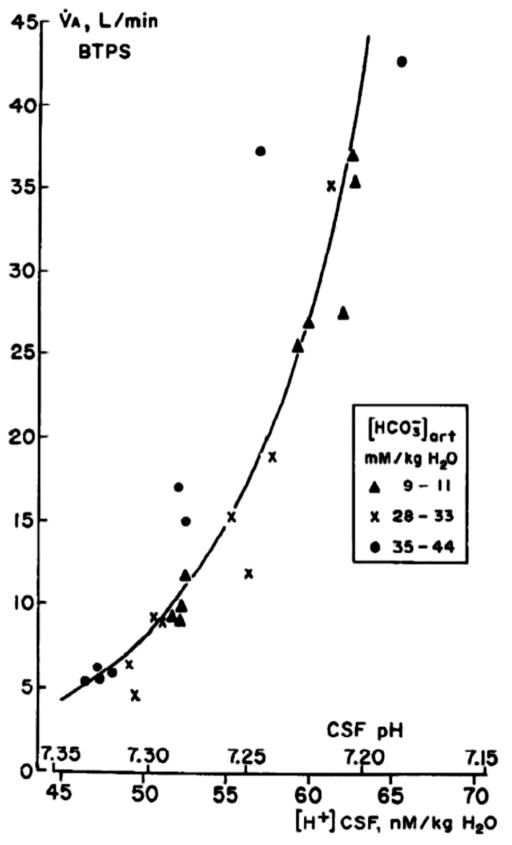
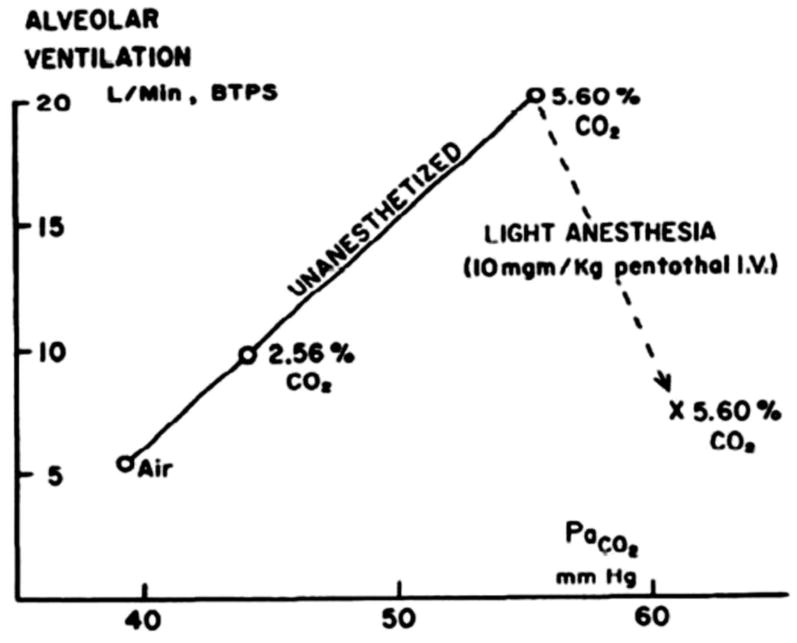
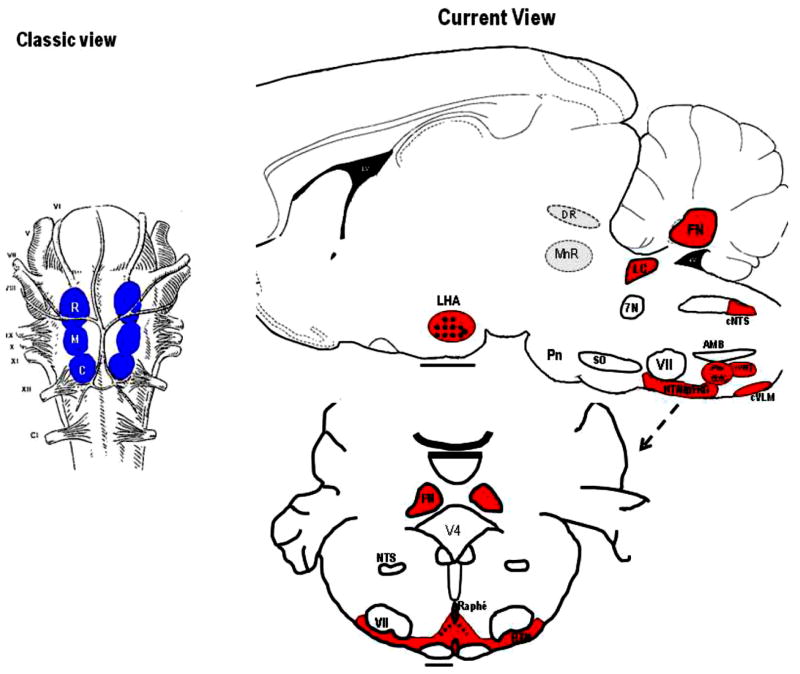
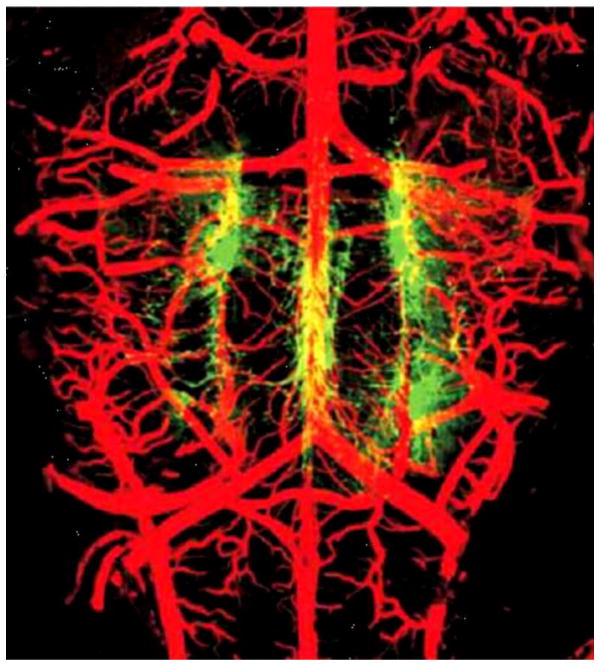
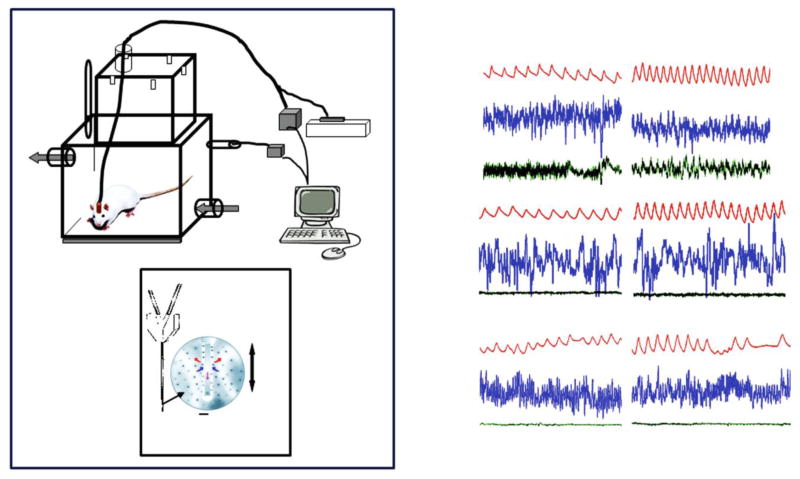
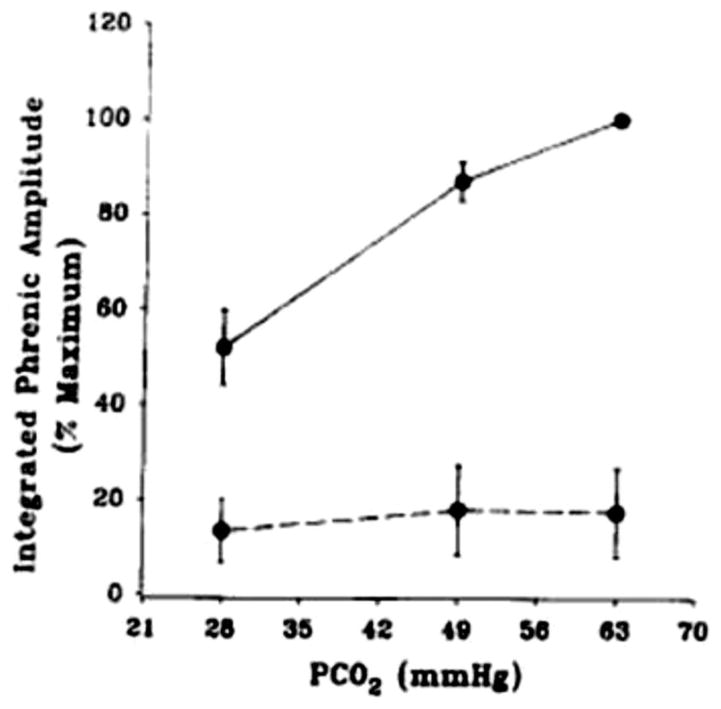
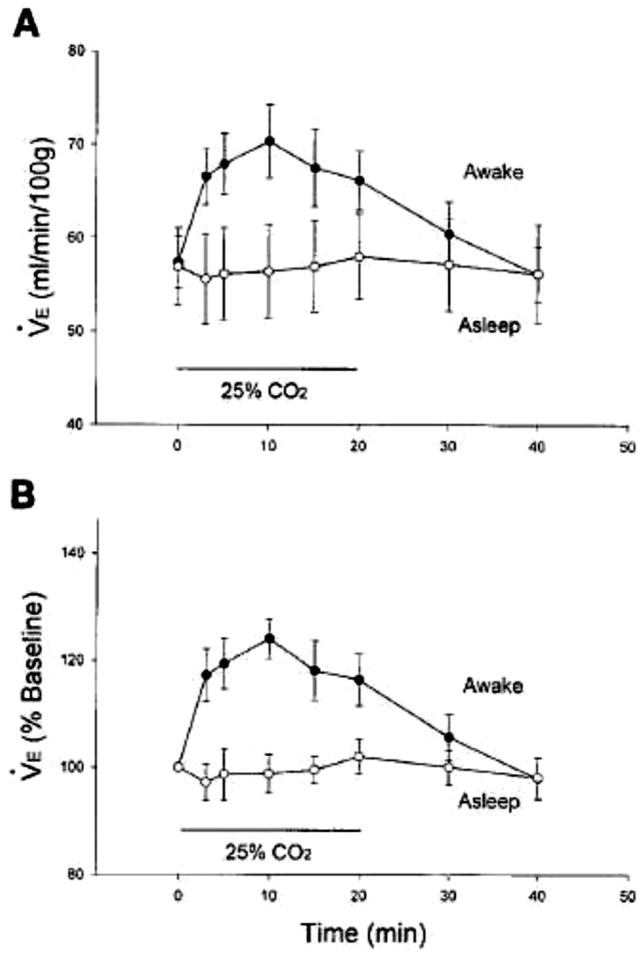
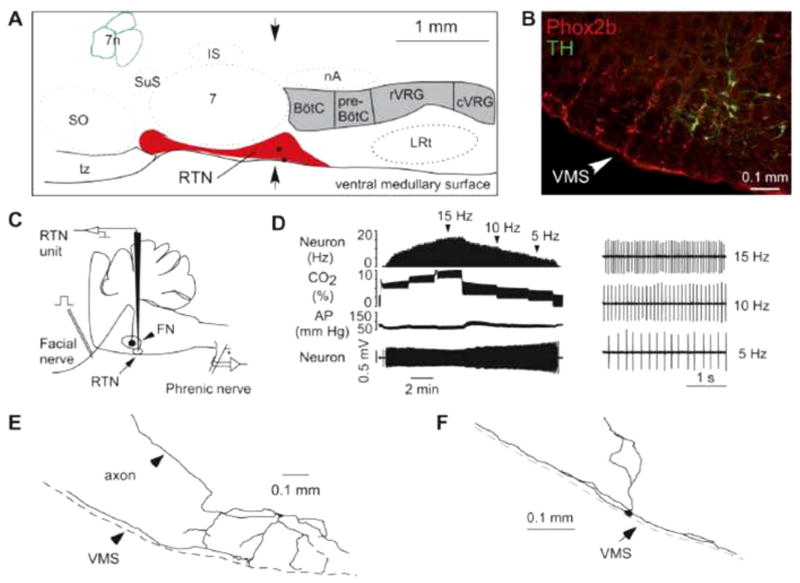
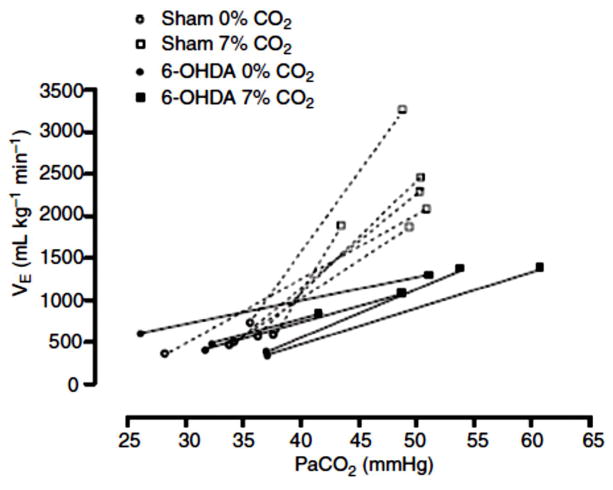
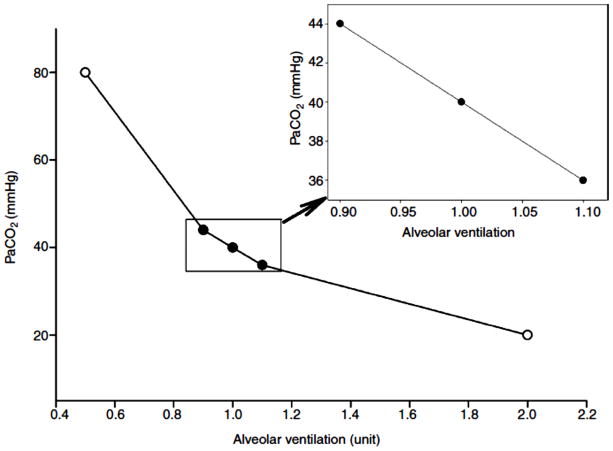
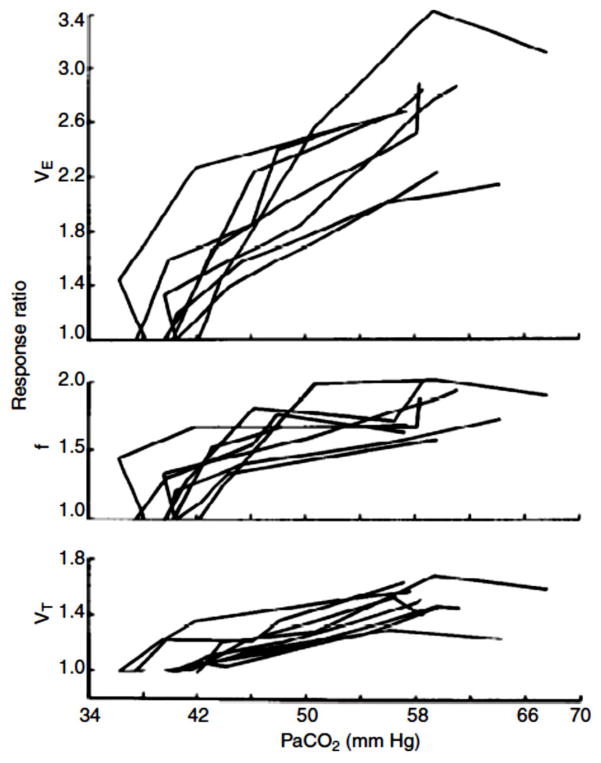
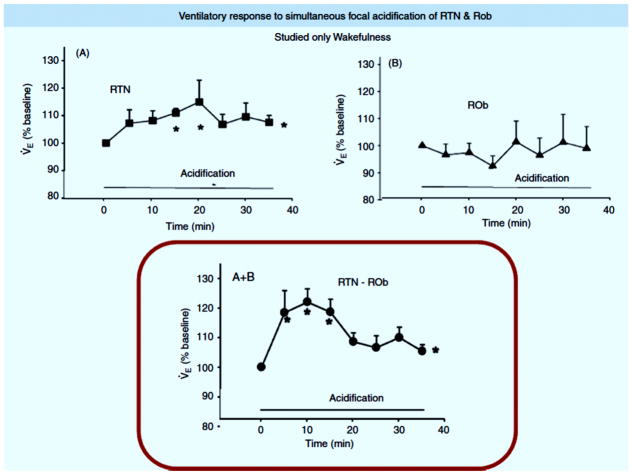
References
-
- Ainslie PN, Duffin J. Integration of cerebrovascular CO2 reactivity and chemoreflex control of breathing: Mechanisms of regulation, measurement, and interpretation. Am J Physiol. 2009;296:R1473–R1495. - PubMed
-
- Akilesh MR, Kamper M, Li A, Nattie EE. Effects of unilateral lesions of retrotrapezoid nucleus on breathing in awake rats. J Appl Physiol. 1997;82:469–479. - PubMed
-
- Amiel J, Dubreuil V, Ramanantsoa N, Fortin G, Gallego J, Brunet JF, Goridis C. PHOX2B in respiratory control: Lessons from congenital central hypoventilation syndrome and its mousemodels. Respir Physiol Neurobiol. 2009;168:125–132. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
