

Alcohol-induced IL-1β in the brain is mediated by NLRP3/ASC inflammasome activation that amplifies neuroinflammation
- PMID: 23625200
- PMCID: PMC3685015
- DOI: 10.1189/jlb.1212659
Alcohol-induced IL-1β in the brain is mediated by NLRP3/ASC inflammasome activation that amplifies neuroinflammation
Abstract
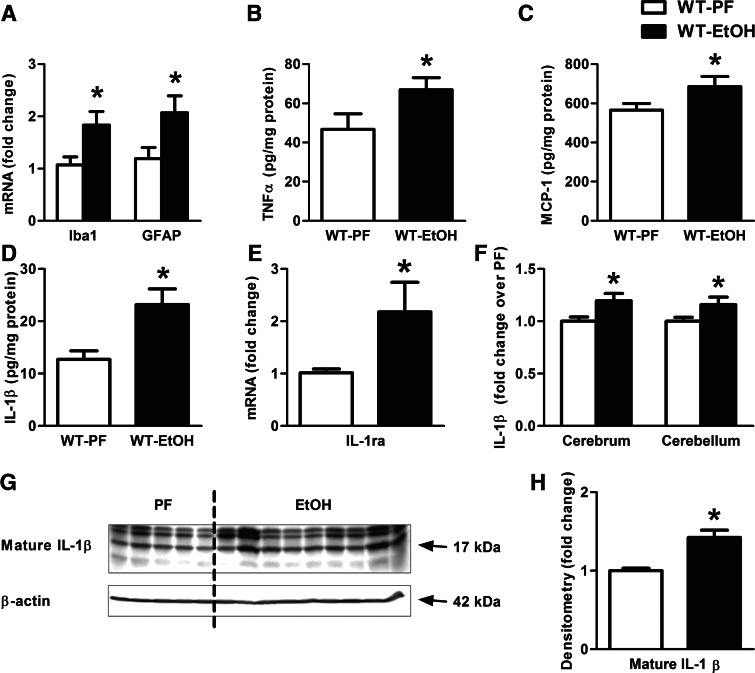
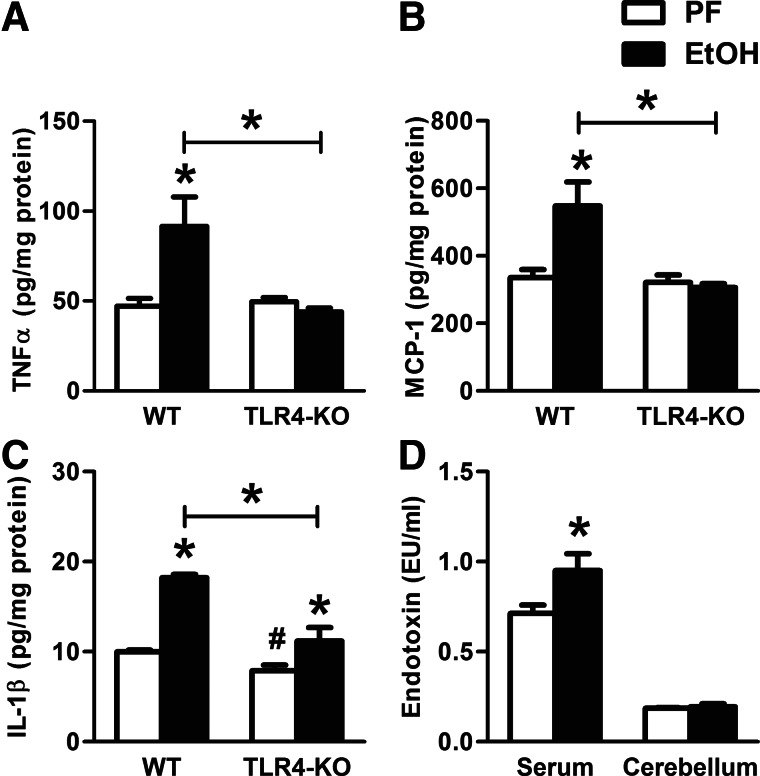
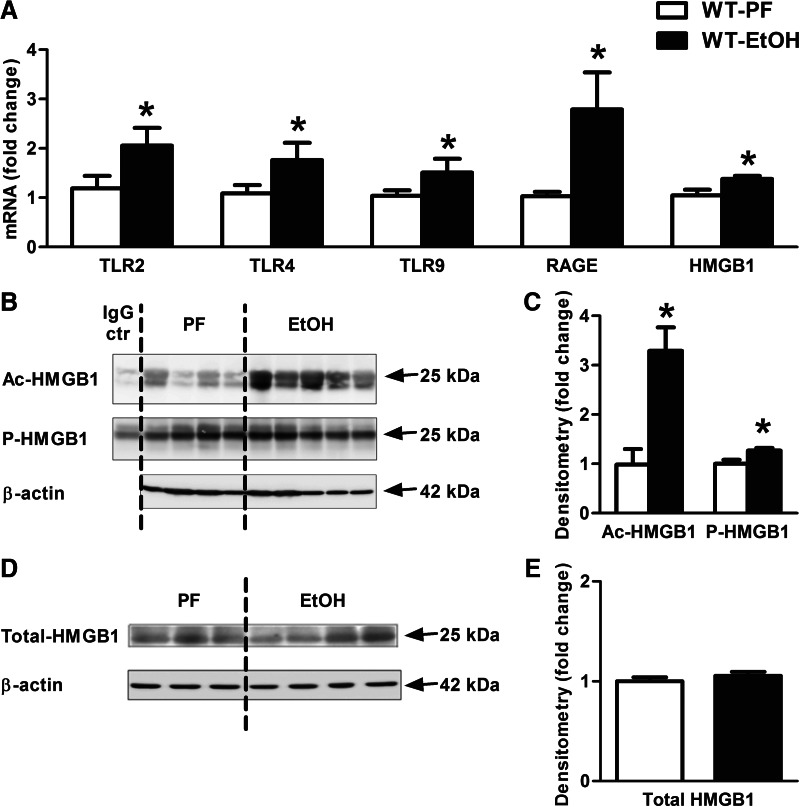
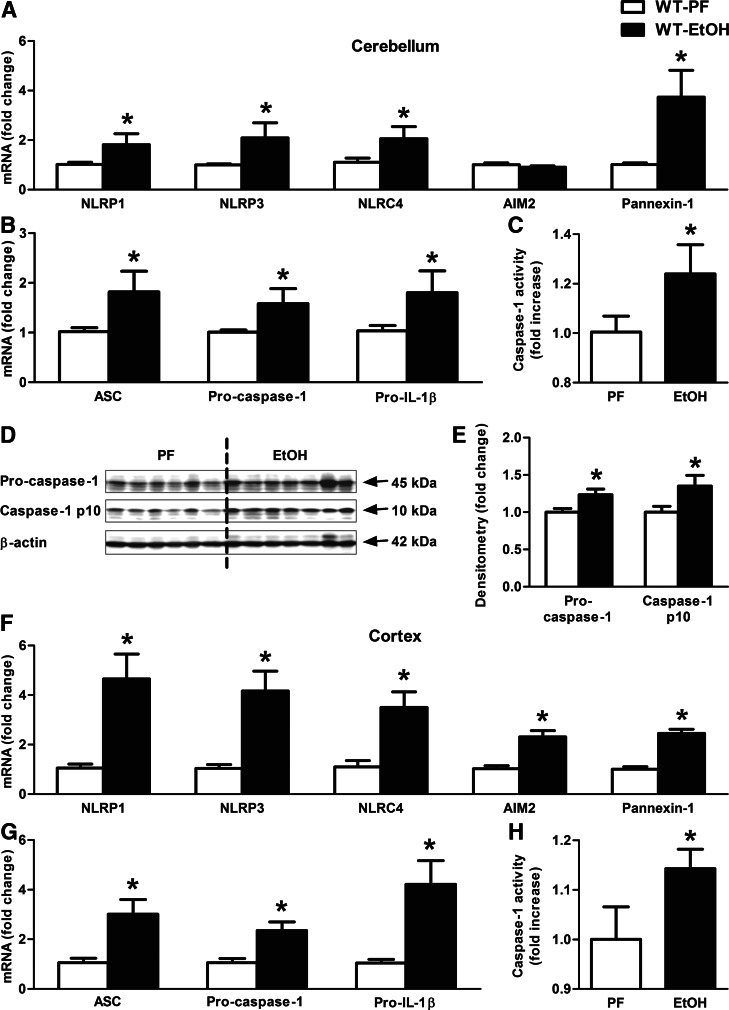
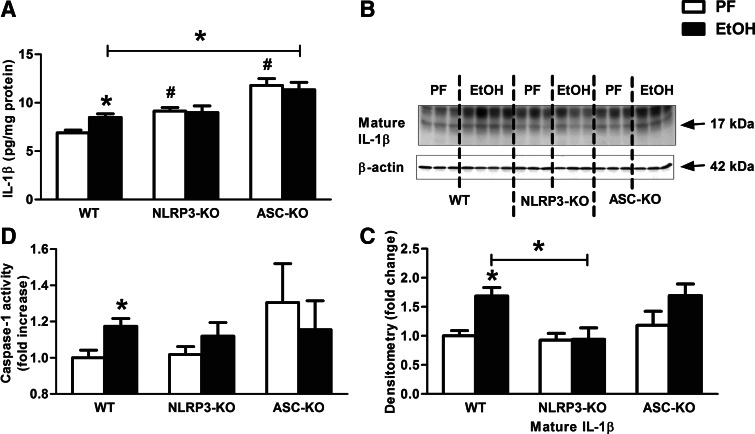
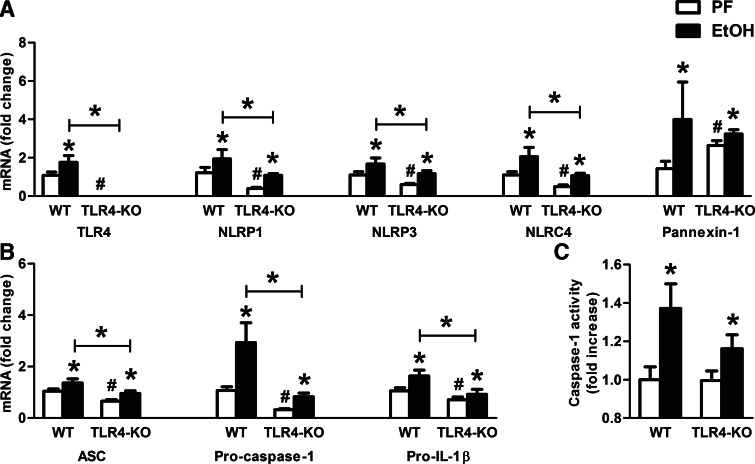
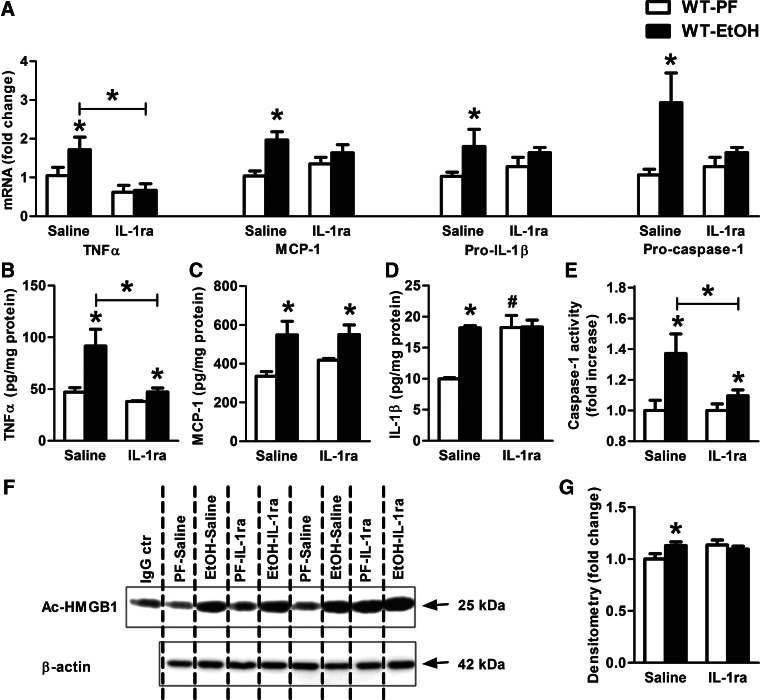
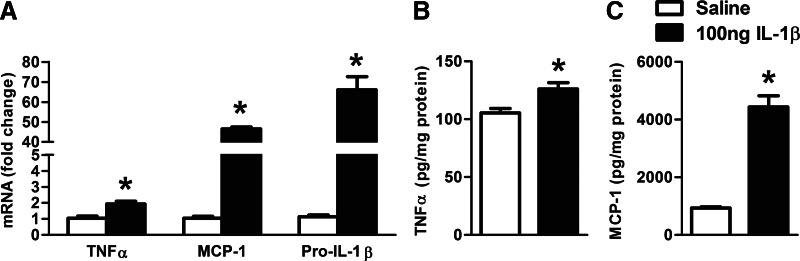
Alcohol-induced neuroinflammation is mediated by proinflammatory cytokines, including IL-1β. IL-1β production requires caspase-1 activation by inflammasomes-multiprotein complexes that are assembled in response to danger signals. We hypothesized that alcohol-induced inflammasome activation contributes to increased IL-1β in the brain. WT and TLR4-, NLRP3-, and ASC-deficient (KO) mice received an ethanol-containing or isocaloric control diet for 5 weeks, and some received the rIL-1ra, anakinra, or saline treatment. Inflammasome activation, proinflammatory cytokines, endotoxin, and HMGB1 were measured in the cerebellum. Expression of inflammasome components (NLRP1, NLRP3, ASC) and proinflammatory cytokines (TNF-α, MCP-1) was increased in brains of alcohol-fed compared with control mice. Increased caspase-1 activity and IL-1β protein in ethanol-fed mice indicated inflammasome activation. TLR4 deficiency protected from TNF-α, MCP-1, and attenuated alcohol-induced IL-1β increases. The TLR4 ligand, LPS, was not increased in the cerebellum. However, we found up-regulation of acetylated and phosphorylated HMGB1 and increased expression of the HMGB1 receptors (TLR2, TLR4, TLR9, RAGE) in alcohol-fed mice. NLRP3- or ASC-deficient mice were protected from caspase-1 activation and alcohol-induced IL-1β increase in the brain. Furthermore, in vivo treatment with rIL-1ra prevented alcohol-induced inflammasome activation and IL-1β, TNF-α, and acetylated HMGB1 increases in the cerebellum. Conversely, intracranial IL-1β administration induced TNF-α and MCP-1 in the cerebellum. In conclusion, alcohol up-regulates and activates the NLRP3/ASC inflammasome, leading to caspase-1 activation and IL-1β increase in the cerebellum. IL-1β amplifies neuroinflammation, and disruption of IL-1/IL-1R signaling prevents alcohol-induced inflammasome activation and neuroinflammation. Increased levels of acetylated and phosphorylated HMGB1 may contribute to alcoholic neuroinflammation.
Keywords: CNS; HMGB1; MCP-1; TNF-α; cerebellum.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
