

Polyamine pathway contributes to the pathogenesis of Parkinson disease
- PMID: 20837543
- PMCID: PMC2947879
- DOI: 10.1073/pnas.1011751107
Polyamine pathway contributes to the pathogenesis of Parkinson disease
Abstract
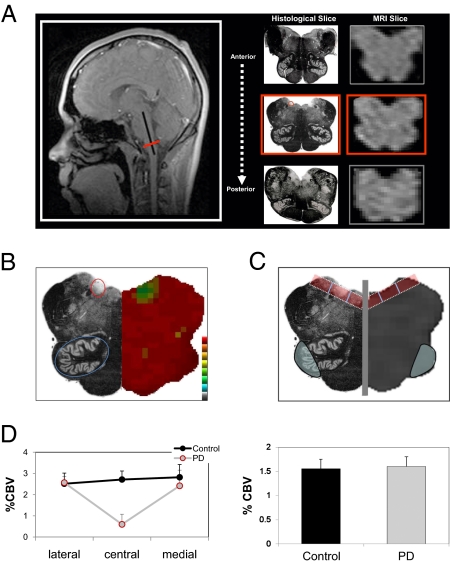
The full complement of molecular pathways contributing to the pathogenesis of Parkinson disease (PD) remains unknown. Here we address this issue by taking a broad approach, beginning by using functional MRI to identify brainstem regions differentially affected and resistant to the disease. Relying on these imaging findings, we then profiled gene expression levels from postmortem brainstem regions, identifying a disease-related decrease in the expression of the catabolic polyamine enzyme spermidine/spermine N1-acetyltransferase 1 (SAT1). Next, a range of studies were completed to support the pathogenicity of this finding. First, to test for a causal link between polyamines and α-synuclein toxicity, we investigated a yeast model expressing α-synuclein. Polyamines were found to enhance the toxicity of α-synuclein, and an unbiased genome-wide screen for modifiers of α-synuclein toxicity identified Tpo4, a member of a family of proteins responsible for polyamine transport. Second, to test for a causal link between SAT1 activity and PD histopathology, we investigated a mouse model expressing α-synuclein. DENSPM (N1, N11-diethylnorspermine), a polyamine analog that increases SAT1 activity, was found to reduce PD histopathology, whereas Berenil (diminazene aceturate), a pharmacological agent that reduces SAT1 activity, worsened the histopathology. Third, to test for a genetic link, we sequenced the SAT1 gene and a rare but unique disease-associated variant was identified. Taken together, the findings from human patients, yeast, and a mouse model implicate the polyamine pathway in PD pathogenesis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Comment in
-
Neurodegenerative disease: Pieces of the Parkinson's puzzle.Nat Rev Neurosci. 2010 Dec;11(12):787. doi: 10.1038/nrn2945. Nat Rev Neurosci. 2010. PMID: 21132875 No abstract available.
References
-
- Hardy J, Cai H, Cookson MR, Gwinn-Hardy K, Singleton A. Genetics of Parkinson's disease and parkinsonism. Ann Neurol. 2006;60:389–398. - PubMed
-
- Litvan I, et al. The etiopathogenesis of Parkinson disease and suggestions for future research. Part I. J Neuropathol Exp Neurol. 2007;66:251–257. - PubMed
-
- Coppola G, Geschwind DH. Technology insight: Querying the genome with microarrays—progress and hope for neurological disease. Nat Clin Pract Neurol. 2006;2:147–158. - PubMed
-
- Kerr MK, Martin M, Churchill GA. Analysis of variance for gene expression microarray data. J Comput Biol. 2000;7:819–837. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
