

Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1
- PMID: 19124468
- PMCID: PMC2649108
- DOI: 10.1074/jbc.M806599200
Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1
Abstract
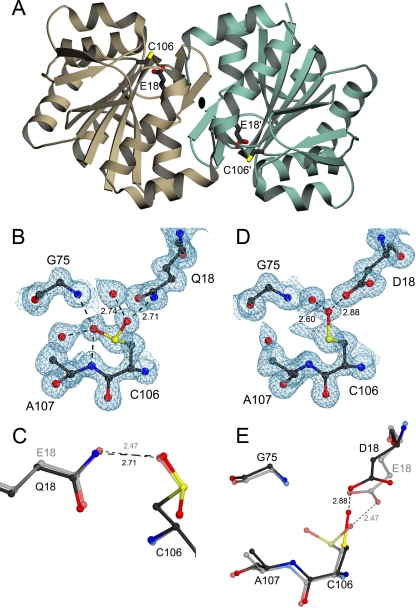
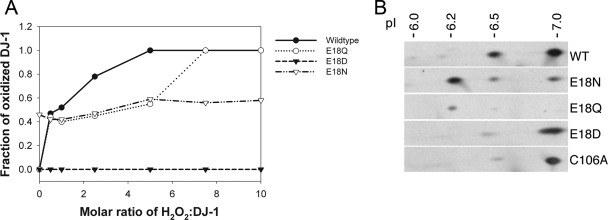
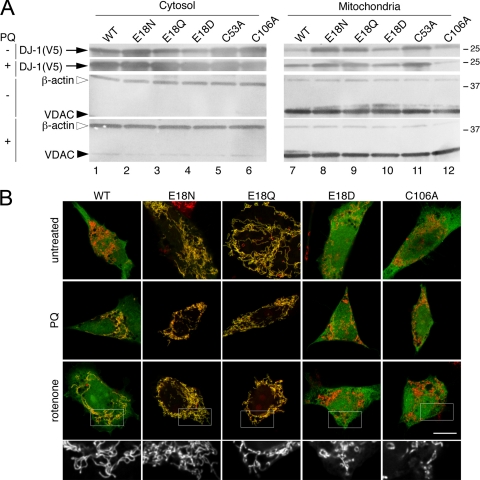
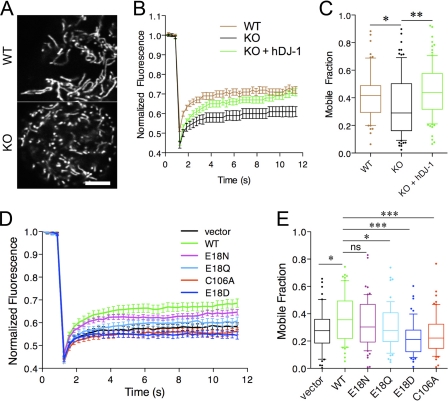
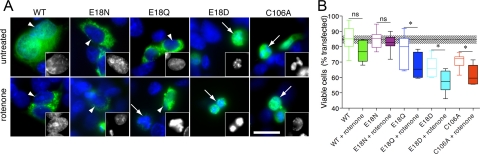
The formation of cysteine-sulfinic acid has recently become appreciated as a modification that links protein function to cellular oxidative status. Human DJ-1, a protein associated with inherited parkinsonism, readily forms cysteine-sulfinic acid at a conserved cysteine residue (Cys106 in human DJ-1). Mutation of Cys106 causes the protein to lose its normal protective function in cell culture and model organisms. However, it is unknown whether the loss of DJ-1 protective function in these mutants is due to the absence of Cys106 oxidation or the absence of the cysteine residue itself. To address this question, we designed a series of substitutions at a proximal glutamic acid residue (Glu18) in human DJ-1 that alter the oxidative propensity of Cys106 through changes in hydrogen bonding. We show that two mutations, E18N and E18Q, allow Cys106 to be oxidized to Cys106-sulfinic acid under mild conditions. In contrast, the E18D mutation stabilizes a cysteine-sulfenic acid that is readily reduced to the thiol in solution and in vivo. We show that E18N and E18Q can both partially substitute for wild-type DJ-1 using mitochondrial fission and cell viability assays. In contrast, the oxidatively impaired E18D mutant behaves as an inactive C106A mutant and fails to protect cells. We therefore conclude that formation of Cys106-sulfinic acid is a key modification that regulates the protective function of DJ-1.
Figures
References
-
- Reddie, K. G., and Carroll, K. S. (2008) Curr. Opin. Chem. Biol. 12 746-754 - PubMed
-
- Biteau, B., Labarre, J., and Toledano, M. B. (2003) Nature 425 980-984 - PubMed
-
- Annesi, G., Savettieri, G., Pugliese, P., D'Amelio, M., Tarantino, P., Ragonese, P., La Bella, V., Piccoli, T., Civitelli, D., Annesi, F., Fierro, B., Piccoli, F., Arabia, G., Caracciolo, M., Cirò Candiano, I. C., and Quattrone, A. (2005) Ann. Neurol. 58 803-807 - PubMed
-
- Bonifati, V., Rizzu, P., van Baren, M. J., Schaap, O., Breedveld, G. J., Krieger, E., Dekker, M. C., Squitieri, F., Ibanez, P., Joosse, M., van Dongen, J. W., Vanacore, N., van Swieten, J. C., Brice, A., Meco, G., van Duijn, C. M., Oostra, B. A., and Heutink, P. (2003) Science 299 256-259 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
