

Early response of alpha2(I) collagen to acetaldehyde in human hepatic stellate cells is TGF-beta independent
- PMID: 16025520
- PMCID: PMC1314984
- DOI: 10.1002/hep.20798
Early response of alpha2(I) collagen to acetaldehyde in human hepatic stellate cells is TGF-beta independent
Abstract
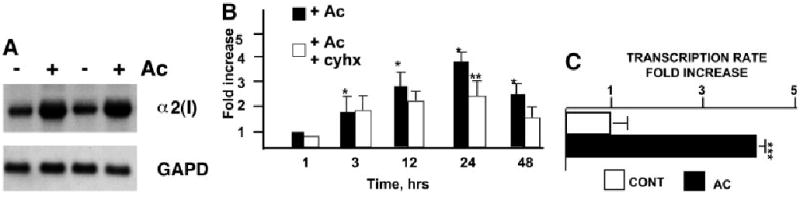
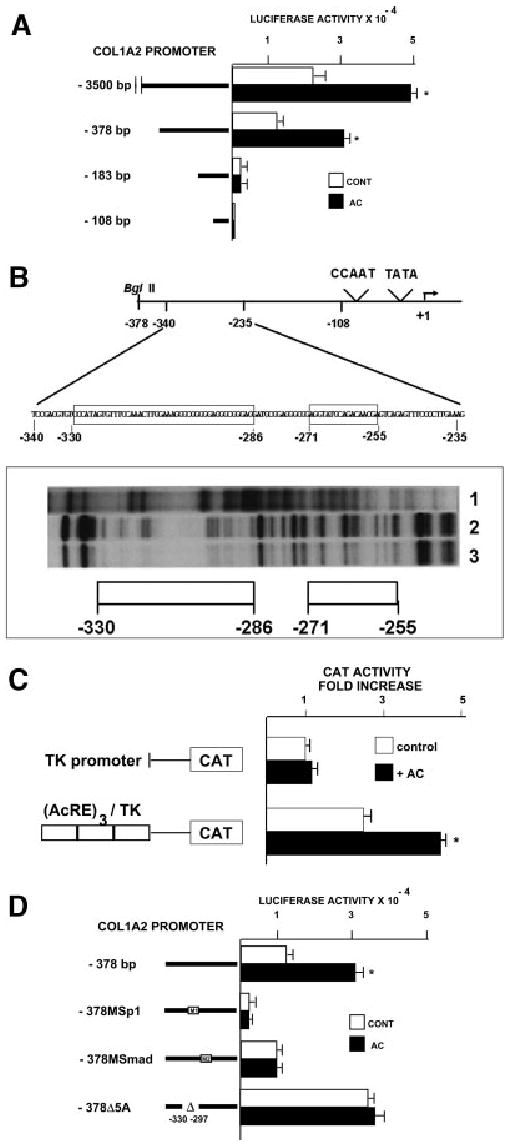
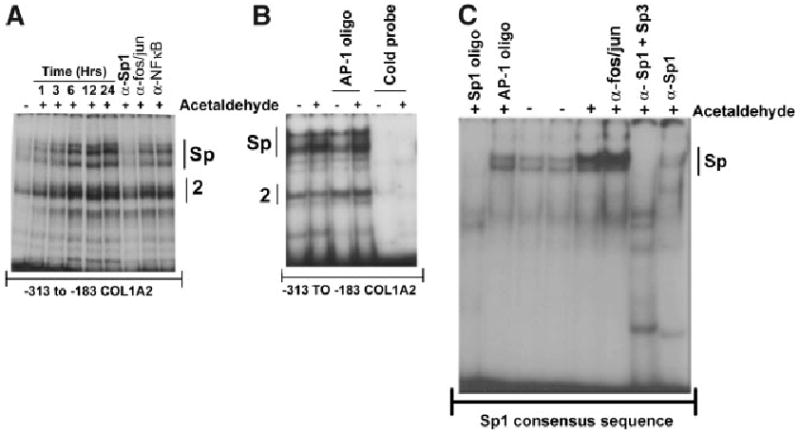
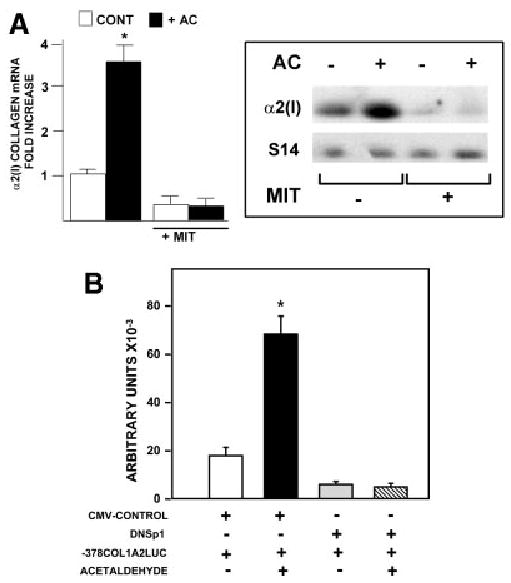
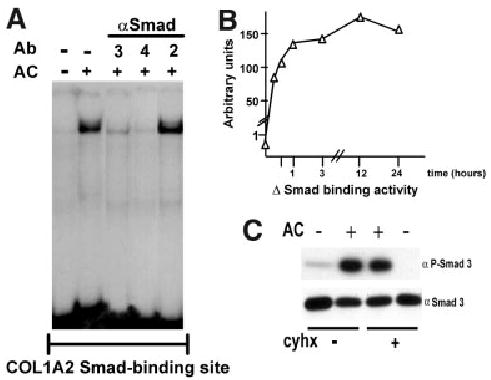
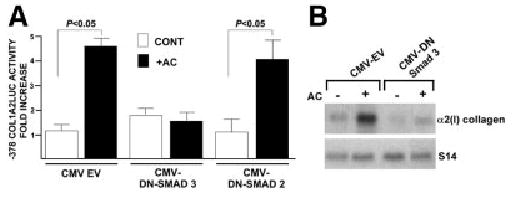
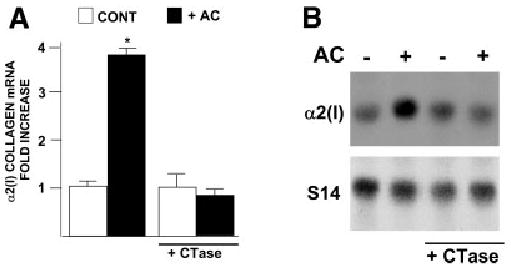
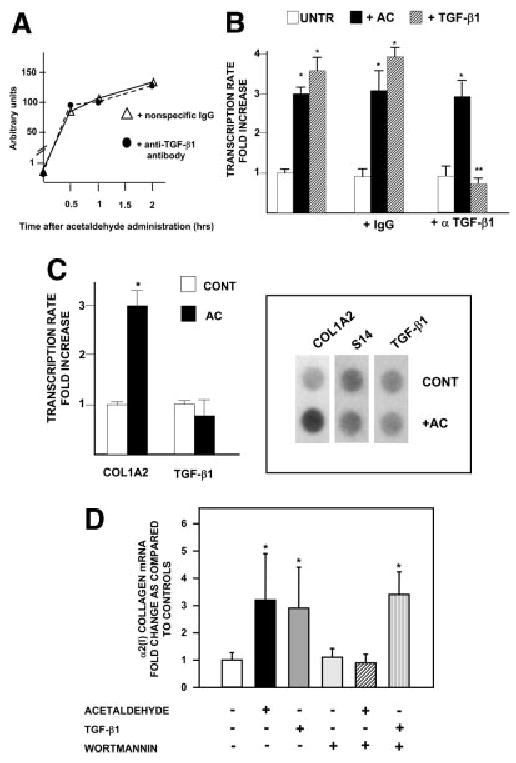
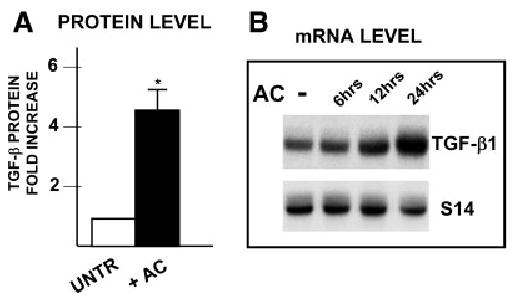
Acetaldehyde is fibrogenic and induces the expression of type I collagen genes in hepatic stellate cells. Some of these acetaldehyde-dependent events are mediated by H(2)O(2) and thus establish a direct connection between oxidative stress and collagen upregulation. We localized to the -378 to -183 region of the alpha2(I) collagen (COL1A2) promoter an acetaldehyde-responsive element (AcRE) functional in human hepatic stellate cells (HHSCs) and investigated molecular mechanisms whereby acetaldehyde stimulates and modulates its transcriptional activity. Because the AcRE co-localized with a previously described transforming growth factor beta (TGF-beta)1-responsive element, and both acetaldehyde and this cytokine induce their effects through H(2)O(2), we investigated whether all fibrogenic actions of acetaldehyde were mediated by this cytokine. Here we show that acetaldehyde-induced COL1A2 upregulation in HHSCs recognizes two distinct but overlapping early and late stages that last from 1 to 6 hours and from 6 to 24 hours, respectively. We present several lines of evidence to show that early acetaldehyde-mediated events are independent of TGF-beta1. These include significant time-course differences in the expression of COL1A2 and TGF-beta1 mRNAs and inability of neutralizing antibodies to TGF-beta1 to inhibit acetaldehyde-dependent collagen gene transcription and Smad 3 phosphorylation. We also show that although acetaldehyde-dependent upregulation of collagen was PI3K dependent, that of TGF-beta1 was PI3K independent. In conclusion, acetaldehyde-dependent mechanisms involved in COL1A2 upregulation are similar, but not identical, to those of TGF-beta1. We suggest that early acetaldehyde-dependent events induce the late expression of TGF-beta1 and create an H(2)O(2)-dependent autocrine loop that may sustain and amplify the fibrogenic response of this alcohol metabolite.
Conflict of interest statement
Potential conflict of interest: Nothing to report.
Figures
References
-
- McClain CJ, Barve S, Deaciuc I, Kugelmas M, Hill D. Cytokines in alcoholic liver disease. Semin Liver Dis. 1999;19:205–219. - PubMed
-
- McClain CJ, Barve S, Barve S, Deaciuc I, Hill DB. Tumor necrosis factor and alcoholic liver disease. Alcohol Clin Exp Res. 1998;22:248S–252S. - PubMed
-
- Hill DB, Marsano L, Cohen D, Allen J, Shedlofsky S, McClain CJ. Increased plasma interleukin-6 concentrations in alcoholic hepatitis. J Lab Clin Med. 1992;119:547–552. - PubMed
-
- Nieto N, Dominguez-Rosales JA, Fontana L, Salazar A, Armendariz-Borunda J, Greenwel P, et al. Rat hepatic stellate cells contribute to the acute-phase response with increased expression of alpha1(I) and alpha1(IV) collagens, tissue inhibitor of metalloproteinase-1, and matrix-metalloproteinase-2 messenger RNAs. Hepatology. 2001;33:597–607. - PubMed
-
- Greenwel P, Rojkind M. Accelerated development of liver fibrosis in CCl4-treated rats by the weekly induction of acute phase response episodes: upregulation of alpha1(I) procollagen and tissue inhibitor of metal-loproteinase-1 mRNAs. Biochim Biophys Acta. 1997;1361:177–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous